The Gastrointestinal Exertional Heat Stroke Paradigm: Pathophysiology, Assessment, Severity, Aetiology and Nutritional Countermeasures

 ,
,
Abstract
:1. Introduction
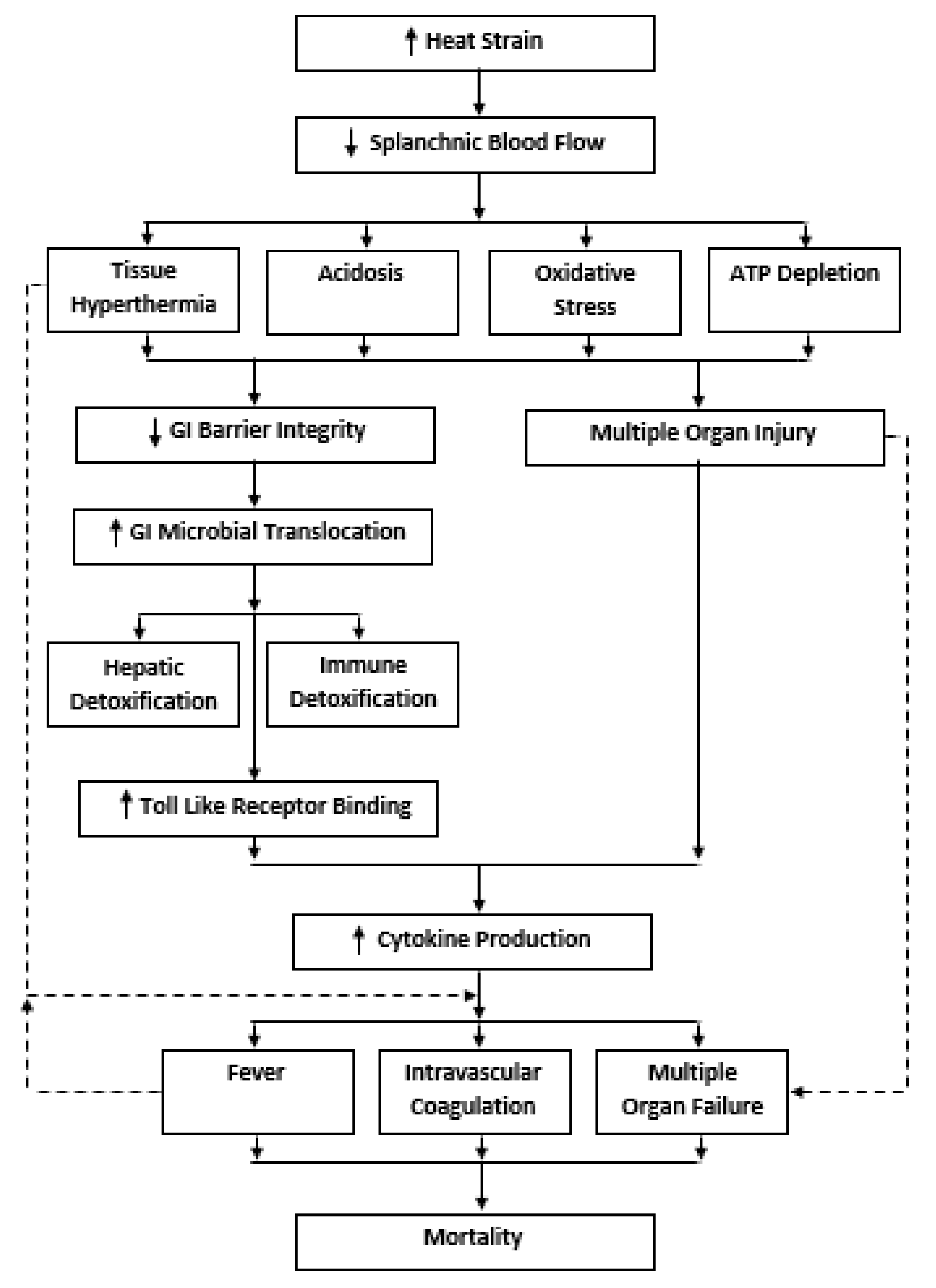
2. The GI Exertional Heat Stroke Paradigm
3. Assessment of GI Barrier Integrity
4. Severity of GI Barrier Integrity Loss Following Exertional-Heat Stress
5. Aetiology of GI Barrier Integrity Loss following Exertional-Heat Stress
6. Nutritional Countermeasures
7. Carbohydrate
8. Glutamine
9. Bovine Colostrum
10. Nitric Oxide
11. Probiotics
12. Polyphenols
13. Zinc-Carnosine
14. Limitations and Future Directions
15. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| BC | Bovine Colostrum |
| Caco-2 | Human Colonic Carcinoma Cell Line |
| CFU | Colony Forming Units |
| CHO | Carbohydrate |
| CHS | Classic Heat Stroke |
| DSAT | Dual-Sugar Absorption Test |
| EHS | Exertional Heat Stroke |
| GI | Gastrointestinal |
| I-BABP | Ileal Bile-Acid Binding Protein |
| I-FABP | Intestinal Fatty Acid Binding Protein |
| IFN | Interferon |
| IGF-1 | Insulin-Like Growth Factor-1 |
| I-HSP | Intracellular Heat Shock Protein |
| IL | Interleukin |
| kDa | Kilodalton |
| LBP | Lipopolysaccharide Binding Protein |
| LPS | Lipopolysaccharide |
| L/R | Lactulose-to-Rhamnose Ratio |
| MOF | Multiple Organ Failure |
| MSAT | Multi-Sugar Absorption Test |
| MT | Microbial Translocation |
| NO | Nitric Oxide |
| NO3 | Nitrate |
| NO2 | Nitrite |
| NOS | Nitric Oxide Synthase |
| NSAIDs | Non-Steroidal Anti-Inflammatory Drugs |
| PAMPs | Pathogen Associated-Molecular Patterns |
| PCT | Procalcitonin |
| RDA | Recommended Daily Allowance |
| RES | Reticuloendothelial system |
| RH | Relative Humidity |
| SAPS | Simplified Acute Physiology Score |
| sCD14 | Soluble Cluster of Differentiation 14 |
| SIRS | Systemic Inflammatory Response Syndrome |
| S/E | Sucralose-to-Erythritol Ratio |
| S/R | Sucrose-to-Rhamnose Ratio |
| Tcore | Core Body Temperature |
| TJ | Tight Junction |
| TLR | Toll-Like Receptor |
| TNF | Tumour Necrosis Factor |
| VO2max | Maximal Oxygen Uptake |
| Wattmax | Maximal Power (wattage) Output |
| ZnC | Zinc Carnosine |
References
- Leon, L.R.; Helwig, B.G. Heat stroke: Role of the systemic inflammatory response. J. Appl. Physiol. 2010, 109, 1980–1988. [Google Scholar] [CrossRef] [PubMed]
- Casa, D.J.; Armstrong, L.E.; Carter, R.; Lopez, R.; Mcdermott, B.; Scriber, K. Historical perspectives on medical care for heat stroke, part 1: Ancient times through the nineteenth century: A review of the literature. Athl. Train. Sports Health Care 2010, 2, 132–138. [Google Scholar] [CrossRef]
- Casa, D.J.; Armstrong, L.E.; Carter, R.; Lopez, R.; Mcdermott, B.; Scriber, K. Historical perspectives on medical care for heat stroke, part 2: 1850 through the present: A review of the literature. Athl. Train. Sports Health Care 2010, 2, 178–190. [Google Scholar] [CrossRef]
- Laitano, O.; Leon, L.R.; Roberts, W.O.; Sawka, M.N. Controversies in exertional heat stroke diagnosis, prevention, and treatment. J. Appl. Physiol. 2019, 127, 1338–1348. [Google Scholar] [CrossRef]
- Bouchama, A.; Knochel, J.P. Heat stroke. N. Eng. J. Med. 2002, 346, 1978–1988. [Google Scholar] [CrossRef]
- Kravchenko, J.; Abernethy, A.P.; Fawzy, M.; Lyerly, H.K. Minimization of heatwave morbidity and mortality. Am. J. Prev. Med. 2013, 44, 274–282. [Google Scholar] [CrossRef]
- Epstein, Y.; Yanovich, R. Heatstroke. N. Engl. J. Med. 2019, 380, 2449–2459. [Google Scholar] [CrossRef]
- Cramer, M.N.; Jay, O. Biophysical aspects of human thermoregulation during heat stress. Auton. Neurosci. 2016, 196, 3–13. [Google Scholar] [CrossRef]
- DeGroot, D.W.; Mok, G.; Hathaway, N.E. International Classification of Disease coding of exertional heat illness in US Army Soldiers. Mil. Med. 2017, 182, e1946–e1950. [Google Scholar] [CrossRef] [Green Version]
- Stacey, M.J.; Parsons, I.T.; Woods, D.R.; Taylor, P.N.; Ross, D.; Brett, S.J. Susceptibility to exertional heat illness and hospitalisation risk in UK military personnel. BMJ Open Sport Exerc. Med. 2015, 1, e000055. [Google Scholar] [CrossRef] [Green Version]
- Kerr, Z.Y.; Casa, D.J.; Marshall, S.W.; Comstock, R.D. Epidemiology of exertional heat illness among US high school athletes. Am. J. Prev. Med. 2013, 44, 8–14. [Google Scholar] [CrossRef] [PubMed]
- Military Health System. Available online: https://health.mil/News/Articles/2019/04/01/Update-Heat-Illness (accessed on 20 January 2020).
- Yeargin, S.W.; Kerr, Z.Y.; Casa, D.J.; Djoko, A.; Hayden, R.; Parsons, J.T.; Dompier, T.P. Epidemiology of Exertional Heat Illnesses in Youth, High School, and College Football. Med. Sci. Sports Exerc. 2016, 48, 1523–1529. [Google Scholar] [CrossRef] [PubMed]
- Yeargin, S.W.; Dompier, T.P.; Casa, D.J.; Hirschhorn, R.M.; Kerr, Z.Y. Epidemiology of Exertional Heat Illnesses in National Collegiate Athletic Association Athletes During the 2009–2010 Through 2014–2015 Academic Years. J. Athl. Train. 2019, 54, 55–63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DeMartini, J.K.; Casa, D.J.; Belval, L.N.; Crago, A.; Davis, R.J.; Jardine, J.J.; Stearns, R.L. Environmental conditions and the occurrence of exertional heat illnesses and exertional heat stroke at the Falmouth Road Race. J. Athl. Train. 2014, 49, 478–485. [Google Scholar] [CrossRef] [Green Version]
- Divine, J.G.; Daggy, M.W.; Dixon, E.E.; LeBlanc, D.P.; Okragly, R.A.; Hasselfeld, K.A. Case Series of Exertional Heat Stroke in Runners During Early Spring: 2014 to 2016 Cincinnati Flying Pig Marathon. Curr. Sports Med. Rep. 2018, 17, 151–158. [Google Scholar] [CrossRef]
- Hosokawa, Y.; Adams, W.M.; Belval, L.N.; Davis, R.J.; Huggins, R.A.; Jardine, J.L.F.; Katch, R.K.; Stearns, R.L.; Casa, D.J. Exertional heat illness incidence and on-site medical team preparedness in warm weather. Int. J. Biometeorol. 2018, 62, 1147–1153. [Google Scholar] [CrossRef]
- Stacey, M.J.; Brett, S.; Woods, D.; Jackson, S.; Ross, D. Case ascertainment of heat illness in the British Army: Evidence of under-reporting from analysis of Medical and Command notifications, 2009–2013. J. R. Army Med. Corps 2016, 162, 428–433. [Google Scholar] [CrossRef] [Green Version]
- Nelson, D.A.; Deuster, P.A.; O’Connor, F.G.; Kurina, L.M. Timing and predictors of mild and severe heat illness among new military enlistees. Med. Sci. Sports Exerc. 2018, 50, 1603. [Google Scholar] [CrossRef]
- Lucas, R.A.; Epstein, Y.; Kjellstrom, T. Excessive occupational heat exposure: A significant ergonomic challenge and health risk for current and future workers. Extreme Physiol. Med. 2014, 3, 14. [Google Scholar] [CrossRef] [Green Version]
- Belval, L.N.; Casa, D.J.; Adams, W.M.; Chiampas, G.T.; Holschen, J.C.; Hosokawa, Y.; Jardine, J.; Kane, S.F.; Labotz, M.; Lemieux, R.S.; et al. Consensus statement-prehospital care of exertional heat stroke. Prehosp. Emerg. Care 2018, 22, 392–397. [Google Scholar] [CrossRef]
- Hosokawa, Y.; Stearns, R.L.; Casa, D.J. Is Heat Intolerance State or Trait? Sports Med. 2019, 49, 365–370. [Google Scholar] [CrossRef] [PubMed]
- Royburt, M.; Epstein, Y.; Solomon, Z.; Shemer, J. Long-term psychological and physiological effects of heat stroke. Physiol. Behav. 1993, 54, 265–267. [Google Scholar] [CrossRef]
- Wang, J.C.; Chien, W.C.; Chu, P.; Chung, C.H.; Lin, C.Y.; Tsai, S.H. The association between heat stroke and subsequent cardiovascular diseases. PLoS ONE 2019, 14, e0211386. [Google Scholar] [CrossRef] [PubMed]
- Wallace, R.F.; Kriebel, D.; Punnett, L.; Wegman, D.H.; Amoroso, P.J. Prior heat illness hospitalization and risk of early death. Environ. Res. 2007, 104, 290–295. [Google Scholar] [CrossRef] [PubMed]
- Epstein, Y.; Druyan, A.; Heled, Y. Heat injury prevention—A military perspective. J. Strength Cond. Res. 2012, 26, S82–S86. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, K.M.; Cheuvront, S.N.; King, M.A.; Mayer, T.A.; Leon, L.R.; Kenefick, R.W. Use of the heat tolerance test to assess recovery from exertional heat stroke. Temperature 2019, 6, 106–119. [Google Scholar] [CrossRef] [PubMed]
- Bonauto, D.; Anderson, R.; Rauser, E.; Burke, B. Occupational heat illness in Washington State, 1995–2005. Am. J. Ind. Med. 2007, 50, 940–950. [Google Scholar] [CrossRef]
- DeGroot, D.W.; Kenefick, R.W.; Sawka, M.N. Impact of arm immersion cooling during ranger training on exertional heat illness and treatment costs. Mil. Med. 2015, 180, 1178–1183. [Google Scholar] [CrossRef] [Green Version]
- Porter, A.M. The death of a British officer-cadet from heat illness. Lancet 2000, 355, 569–571. [Google Scholar] [CrossRef]
- Stacey, M.; Woods, D.; Ross, D.; Wilson, D. Heat illness in military populations: Asking the right questions for research. J. R. Army Med. Corps 2014, 160, 121–124. [Google Scholar] [CrossRef]
- Armstrong, L.E.; Casa, D.J.; Millard-Stafford, M.; Moran, D.S.; Pyne, S.W.; Roberts, W.O. American College of Sports Medicine position stand. Exertional heat illness during training and competition. Med. Sci. Sports Exerc. 2007, 39, 556–572. [Google Scholar] [CrossRef] [PubMed]
- Casa, D.J.; DeMartini, J.K.; Bergeron, M.F.; Csillan, D.; Eichner, E.R.; Lopez, R.M.; Ferrara, M.S.; Miller, K.C.; O’Connor, F.; Sawka, M.N.; et al. National Athletic Trainers’ Association position statement: Exertional heat illnesses. J. Athl. Train. 2015, 50, 986–1000. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Altman, J.; Stern, E.; Stern, M.; Prine, B.; Smith, K.B.; Smith, M.S. Current paradigms in the prehospital care of exertional heat illness: A review. Curr. Orthop. Pract. 2019, 31. [Google Scholar] [CrossRef]
- Ministry of Defence. Available online: https://assets.publishing.service.gov.uk/government/uploads/system/uploads/attachment_data/file/793094/JSP_539_Part_2_V3.1__Updated_04-19_.pdf (accessed on 20 January 2020).
- Armstrong, L.E.; Lee, E.C.; Armstrong, E.M. Interactions of Gut Microbiota, Endotoxemia, Immune Function, and Diet in Exertional Heatstroke. J. Sports Med. 2018, 2018, 5724575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lim, C.L. Heat sepsis precedes heat toxicity in the pathophysiology of heat stroke—A new paradigm on an ancient disease. Antioxidants 2018, 7, 149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thursby, E.; Juge, N. Introduction to the human gut microbiota. Biochem. J. 2017, 474, 1823–1836. [Google Scholar] [CrossRef]
- Sender, R.; Fuchs, S.; Milo, R. Revised estimates for the number of human and bacteria cells in the body. PLoS Biol. 2016, 14, e1002533. [Google Scholar] [CrossRef] [Green Version]
- Wells, J.M.; Brummer, R.J.; Derrien, M.; MacDonald, T.T.; Troost, F.; Cani, P.D.; Theodorou, V.; Dekker, J.; Méheust, A.; De Vos, W.M.; et al. Homeostasis of the gut barrier and potential biomarkers. Am. J. Physiol. Gastrointest. Liver Physiol. 2016, 312, G171–G193. [Google Scholar] [CrossRef]
- Moseley, P.L.; Gisolfi, C.V. New frontiers in thermoregulation and exercise. Sports Med. 1993, 16, 163–167. [Google Scholar] [CrossRef]
- Munford, R.S. Invited review: Detoxifying endotoxin: Time, place and person. J. Endotoxin Res. 2005, 11, 69–84. [Google Scholar] [CrossRef]
- Deitch, E.A. Gut-origin sepsis: Evolution of a concept. Surgeon 2012, 10, 350–356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gathiram, P.; Gaffin, S.L.; Brock-Utne, J.G.; Wells, M.T. Time course of endotoxemia and cardiovascular changes in heat-stressed primates. Aviation Space Environ. Med. 1987, 58, 1071–1074. [Google Scholar]
- Nieman, D.C.; Nehlsen-Cannarella, S.L. The effects of acute and chronic exercise on immunoglobulins. Sports Med. 1991, 11, 183–201. [Google Scholar] [CrossRef] [PubMed]
- Fukui, H. Endotoxin and other microbial translocation markers in the blood: A clue to understand leaky gut syndrome. Cell Mol Med 2016, 2, 3. [Google Scholar] [CrossRef]
- Heled, Y.; Fleischmann, C.; Epstein, Y. Cytokines and their role in hyperthermia and heat stroke. J. Basic Clin. Physiol. Pharmacol. 2013, 24, 85–96. [Google Scholar] [CrossRef]
- Lim, C.L.; Mackinnon, L.T. The roles of exercise-induced immune system disturbances in the pathology of heat stroke. Sports Med. 2006, 36, 39–64. [Google Scholar] [CrossRef]
- Bynum, G.; Brown, J.; Dubose, D.; Marsili, M.; Leav, I.; Pistole, T.G.; Hamlet, M.; LeMaire, M.; Caleb, B. Increased survival in experimental dog heatstroke after reduction of gut flora. Aviation Space Environ. Med. 1979, 50, 816–819. [Google Scholar]
- Gathiram, P.; Wells, M.T.; Brock-Utne, J.G.; Wessels, B.C.; Gaffin, S.L. Prevention of endotoxaemia by non-absorbable antibiotics in heat stress. J. Clin. Pathol. 1987, 40, 1364–1368. [Google Scholar] [CrossRef] [Green Version]
- Gathiram, P.; Wells, M.T.; Brock-Utne, J.G.; Gaffin, S.L. Antilipopolysaccharide improves survival in primates subjected to heat stroke. Circ. Shock 1987, 23, 157–164. [Google Scholar]
- Gathiram, P.; Wells, M.T.; Brock-Utne, J.G.; Gaffin, S.L. Prophylactic corticosteroid increases survival in experimental heat stroke in primates. Aviation Space Environ. Med. 1988, 59, 352–355. [Google Scholar]
- Lim, C.L.; Wilson, G.; Brown, L.; Coombes, J.S.; Mackinnon, L.T. Pre-existing inflammatory state compromises heat tolerance in rats exposed to heat stress. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2007, 292, R186–R194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, T.H.; Shih, M.F.; Wen, Y.S.; Ho, W.Y.; Leu, K.L.; Wang, M.Y.; Liu, C.C. Attenuation of circulatory shock and cerebral ischemia injury in heat stroke by combination treatment with dexamethasone and hydroxyethyl starch. Exp. Transl. Stroke Med. 2010, 2, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, C.C.; Shih, M.F.; Wen, Y.S.; Lai, Y.H.; Yang, T.H. Dexamethasone improves heat stroke-induced multiorgan dysfunction and damage in rats. Int. J. Mol. Sci. 2014, 15, 21299–21313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Audet, G.N.; Dineen, S.M.; Stewart, D.A.; Plamper, M.L.; Pathmasiri, W.W.; McRitchie, S.L.; Sumner, S.J.; Leon, L.R. Pretreatment with indomethacin results in increased heat stroke severity during recovery in a rodent model of heat stroke. J. Appl. Physiol. 2017, 123, 544–557. [Google Scholar] [CrossRef]
- Caputa, M.; Dokladny, K.; Kurowicka, B. Endotoxaemia does not limit heat tolerance in rats: The role of plasma lipoproteins. Eur. J. Appl. Physiol. 2000, 82, 142–150. [Google Scholar] [CrossRef]
- Lin, X.J.; Li, Y.J.; Li, Z.L.; Zou, F.; Lin, M.T. Pre-existing lipopolysaccharide may increase the risk of heatstroke in rats. Am. J. Med. Sci. 2009, 337, 265–270. [Google Scholar] [CrossRef]
- King, M.A.; Leon, L.R.; Mustico, D.L.; Haines, J.M.; Clanton, T.L. Biomarkers of multiorgan injury in a preclinical model of exertional heat stroke. J. Appl. Physiol. 2015, 118, 1207–1220. [Google Scholar] [CrossRef] [Green Version]
- Lin, Y.; Zhang, Y. Renoprotective effect of oral rehydration solution III in exertional heatstroke rats. Renal Fail. 2019, 41, 190–196. [Google Scholar] [CrossRef] [Green Version]
- King, M.A.; Leon, L.R.; Morse, D.A.; Clanton, T.L. Unique cytokine and chemokine responses to exertional heat stroke in mice. J. Appl. Physiol. 2016, 122, 296–306. [Google Scholar] [CrossRef]
- Malamud, N.; Haymaker, W.; Custer, R.P. Heat Stroke. A Clinico-Pathologic Study of 125 Fatal Cases. Mil. Surg. 1946, 99, 397–449. [Google Scholar]
- Shibolet, S.; Coll, R.; Gilat, T.; Sohar, E. Heatstroke: Its clinical picture and mechanism in 36 cases. Q. J. Med. 1967, 36, 525–548. [Google Scholar] [PubMed]
- Chao, T.C.; Sinniah, R.; Pakiam, J.E. Acute heat stroke deaths. Pathology 1981, 13, 145–156. [Google Scholar] [CrossRef] [PubMed]
- Graber, C.D.; Reinhold, R.B.; Breman, J.G.; Harley, R.A.; Hennigar, G.R. Fatal heat stroke: Circulating endotoxin and gram-negative sepsis as complications. Jama 1971, 216, 1195–1196. [Google Scholar] [CrossRef] [PubMed]
- Bouchama, A.; Parhar, R.S.; el-Yazigi, A.; Sheth, K.; al-Sedairy, S. Endotoxemia and release of tumor necrosis factor and interleukin 1 alpha in acute heatstroke. J. Appl. Physiol. 1991, 70, 2640–2644. [Google Scholar] [CrossRef] [PubMed]
- Bouchama, A.; Al-Sedairy, S.; Siddiqui, S.; Shail, E.; Bezeig, M. Elevated pyrogenic cytokines in heatstroke. Chest 1993, 104, 1498–1502. [Google Scholar] [CrossRef] [PubMed]
- Lu, K.C.; Wang, J.Y.; Lin, S.H.; Chu, P.; Lin, Y.F. Role of circulating cytokines and chemokines in exertional heatstroke. Crit. Care Med. 2004, 32, 399–403. [Google Scholar] [CrossRef]
- Hashim, I.A.; Al-Zeer, A.; Al-Shohaib, S.; Al-Ahwal, M.; Shenkin, A. Cytokine changes in patients with heatstroke during pilgrimage to Makkah. Med. Inflamm. 1997, 6, 135–139. [Google Scholar] [CrossRef]
- Camus, G.; Nys, M.; Poortmans, J.R.; Venneman, I.; Monfils, T.; Deby-Dupont, G.; Juchmes-Ferir, A.; Deby, C.; Lamy, M.; Duchateau, J. Endotoxaemia, production of tumour necrosis factor α and polymorphonuclear neutrophil activation following strenuous exercise in humans. Eur. J. Appl. Physiol. Occupat. Physiol. 1998, 79, 62–68. [Google Scholar] [CrossRef]
- Gill, S.K.; Teixeira, A.; Rama, L.; Prestes, J.; Rosado, F.; Hankey, J.; Scheer, V.; Hemmings, K.; Ansley-Robson, P.; Costa, R.J. Circulatory endotoxin concentration and cytokine profile in response to exertional-heat stress during a multi-stage ultra-marathon competition. Exerc. Immunol. Rev. 2015, 21, 114. [Google Scholar]
- Barberio, M.D.; Elmer, D.J.; Laird, R.H.; Lee, K.A.; Gladden, B.; Pascoe, D.D. Systemic LPS and inflammatory response during consecutive days of exercise in heat. Int. J. Sports Med. 2015, 36, 262–270. [Google Scholar] [CrossRef]
- Ng, Q.Y.; Lee, K.W.; Byrne, C.; Ho, T.F.; Lim, C.L. Plasma Endotoxin and Immune Responses During a 21-km Road Race Under a Warm and Humid Environment. Ann. Acad. Med. Singap. 2008, 37, 307–314. [Google Scholar] [PubMed]
- Aibiki, M.; Ohtsubo, S.; Nishiyama, T.; Maekawa, S.; Oka, H.; Dote, K.; Shirakawa, Y. Elevated serum beta-D-glucan level and depressed neutrophil phagocytosis in a heatstroke patient. Resuscitation 2005, 65, 115–117. [Google Scholar] [CrossRef] [PubMed]
- Wacker, C.; Prkno, A.; Brunkhorst, F.M.; Schlattmann, P. Procalcitonin as a diagnostic marker for sepsis: A systematic review and meta-analysis. Lancet Infect. Dis. 2013, 13, 426–435. [Google Scholar] [CrossRef]
- Tong, H.S.; Liu, Y.S.; Wen, Q.; Tang, Y.Q.; Yuan, F.F.; Su, L. Serum procalcitonin predicting mortality in exertional heatstroke. Emerg. Med. J. 2012, 29, 113–117. [Google Scholar] [CrossRef] [PubMed]
- Hausfater, P.; Hurtado, M.; Pease, S.; Juillien, G.; Lvovschi, V.E.; Salehabadi, S.; Lidove, O.; Wolff, M.; Bernard, M.; Chollet-Martin, S.; et al. Is procalcitonin a marker of critical illness in heatstroke? Intensive Care Med. 2008, 34, 1377–1383. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.; Liu, W.; Yu, J.; Jiang, J.; Xu, T.; Zhou, Y. Effect of prealbumin level on mortality in heatstroke patients. Exp. Ther. Med. 2019, 17, 3053–3060. [Google Scholar] [CrossRef] [Green Version]
- Galipeau, H.J.; Verdu, E.F. The complex task of measuring intestinal permeability in basic and clinical science. Neurogastroenterol. Motil. 2016, 28, 957–965. [Google Scholar] [CrossRef]
- Costa, R.J.S.; Snipe, R.M.J.; Kitic, C.M.; Gibson, P.R. Systematic review: Exercise-induced gastrointestinal syndrome—Implications for health and intestinal disease. Aliment. Pharmacol. Ther. 2017, 46, 246–265. [Google Scholar] [CrossRef] [Green Version]
- Pires, W.; Veneroso, C.E.; Wanner, S.P.; Pacheco, D.A.S.; Vaz, G.C.; Amorim, F.T.; Tonoli, C.; Soares, D.D.; Coimbra, C.C. Association Between Exercise-Induced Hyperthermia and Intestinal Permeability: A Systematic Review. Sports Med. 2017, 47, 1389–1403. [Google Scholar] [CrossRef]
- Van Wijck, K.; Lenaerts, K.; Grootjans, J.; Wijnands, K.A.; Poeze, M.; Van Loon, L.J.; Dejong, C.H.; Buurman, W.A. Physiology and pathophysiology of splanchnic hypoperfusion and intestinal injury during exercise: Strategies for evaluation and prevention. Am. J. Physiol. Gastrointest. Liver Physiol. 2012, 303, G155–G168. [Google Scholar] [CrossRef] [Green Version]
- Menzies, I.S.; Zuckerman, M.J.; Nukajam, W.S.; Somasundaram, S.G.; Murphy, B.; Jenkins, A.P.; Crane, R.S.; Gregory, G.G. Geography of intestinal permeability and absorption. Gut 1999, 44, 483–489. [Google Scholar] [CrossRef] [PubMed]
- Bjarnason, I.; Macpherson, A.; Hollander, D. Intestinal permeability: An overview. Gastroenterology 1995, 108, 1566–1581. [Google Scholar] [CrossRef]
- Fleming, S.C.; Duncan, A.; Russell, R.I.; Laker, M.F. Measurement of sugar probes in serum: An alternative to urine measurement in intestinal permeability testing. Clin. Chem. 1996, 42, 445–448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Wijck, K.; Lenaerts, K.; Van Loon, L.J.; Peters, W.H.; Buurman, W.A.; Dejong, C.H. Exercise-induced splanchnic hypoperfusion results in gut dysfunction in healthy men. PLoS ONE 2011, 6, e22366. [Google Scholar] [CrossRef] [Green Version]
- Van Wijck, K.; van Eijk, H.M.; Buurman, W.A.; Dejong, C.H.; Lenaerts, K. Novel analytical approach to a multi-sugar whole gut permeability assay. J. Chromatogr. B 2011, 879, 2794–2801. [Google Scholar] [CrossRef]
- Pugh, J.N.; Impey, S.G.; Doran, D.A.; Fleming, S.C.; Morton, J.P.; Close, G.L. Acute high-intensity interval running increases markers of gastrointestinal damage and permeability but not gastrointestinal symptoms. Appl. Physiol. Nutr. Metab. 2017, 42, 941–947. [Google Scholar] [CrossRef]
- Ogden, H.B.; Fallowfield, J.D.L.; Child, R.B.; Davison, G.; Fleming, S.C.; Edinburgh, R.M.; Delves, S.K.; Millyard, M.; Westwood, C.S.; Layden, J.D. Reliability of Gastrointestinal Barrier Integrity and Microbial Translocation Biomarkers at Rest and Following Exertional Heat Stress. Physiol. Rep. 2020. [Google Scholar]
- Van Wijck, K.; Verlinden, T.J.; van Eijk, H.M.; Dekker, J.; Buurman, W.A.; Dejong, C.H.; Lenaerts, K. Novel multi-sugar assay for site-specific gastrointestinal permeability analysis: A randomized controlled crossover trial. Clin. Nutr. 2013, 32, 245–251. [Google Scholar] [CrossRef]
- Van Wijck, K.; Bessems, B.A.; van Eijk, H.M.H.; Buurman, W.A.; Dejong, C.H.C.; Lenaerts, K. Polyethylene glycol versus dual sugar assay for gastrointestinal permeability analysis: Is it time to choose? Clin. Exp. Gastroenterol. 2012, 5, 139–150. [Google Scholar] [CrossRef] [Green Version]
- Sun, D.L.; Cen, Y.Y.; Li, S.M.; Li, W.M.; Lu, Q.P.; Xu, P.Y. Accuracy of the serum intestinal fatty-acid-binding protein for diagnosis of acute intestinal ischemia: A meta-analysis. Sci. Rep. 2016, 6, 34371. [Google Scholar] [CrossRef]
- Van de Poll, M.C.; Derikx, J.P.; Buurman, W.A.; Peters, W.H.; Roelofs, H.M.; Wigmore, S.J.; Dejong, C.H. Liver manipulation causes hepatocyte injury and precedes systemic inflammation in patients undergoing liver resection. World J. Surg. 2007, 31, 2033–2038. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montagnana, M.; Danese, E.; Lippi, G. Biochemical markers of acute intestinal ischemia: Possibilities and limitations. Ann. Transl. Med. 2018, 6, 341. [Google Scholar] [CrossRef] [PubMed]
- Sturgeon, C.; Fasano, A. Zonulin, a regulator of epithelial and endothelial barrier functions, and its involvement in chronic inflammatory diseases. Tissue Barriers 2016, 4, e1251384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ajamian, M.; Steer, D.; Rosella, G.; Gibson, P.R. Serum zonulin as a marker of intestinal mucosal barrier function: May not be what it seems. PLoS ONE 2019, 14, e0210728. [Google Scholar] [CrossRef]
- Thuijls, G.; Derikx, J.P.; de Haan, J.J.; Grootjans, J.; de Bruïne, A.; Masclee, A.A.; Heineman, E.; Buurman, W.A. Urine-based detection of intestinal tight junction loss. J. Clin. Gastroenterol. 2010, 44, e14–e19. [Google Scholar] [CrossRef] [PubMed]
- Nagpal, R.; Yadav, H. Bacterial translocation from the gut to the distant organs: An overview. Ann. Nutr. Metab. 2017, 71, 11–16. [Google Scholar] [CrossRef]
- Gnauck, A.; Lentle, R.G.; Kruger, M.C. Chasing a ghost?–Issues with the determination of circulating levels of endotoxin in human blood. Crit. Rev. Clin. Lab. Sci. 2016, 53, 197–215. [Google Scholar] [CrossRef]
- Mussap, M.; Noto, A.; Fravega, M.; Fanos, V. Soluble CD14 subtype presepsin (sCD14-ST) and lipopolysaccharide binding protein (LBP) in neonatal sepsis: New clinical and analytical perspectives for two old biomarkers. J. Matern. Fetal Neonat. Med. 2011, 24 (Suppl. 2), 12–14. [Google Scholar] [CrossRef]
- Mierzchala, M.; Krzystek-Korpacka, M.; Gamian, A.; Durek, G. Quantitative indices of dynamics in concentrations of lipopolysaccharide-binding protein (LBP) as prognostic factors in severe sepsis/septic shock patients—Comparison with CRP and procalcitonin. Clin. Biochem. 2011, 44, 357–363. [Google Scholar] [CrossRef]
- Ewaschuk, J.B.; Naylor, J.M.; Zello, G.A. D-lactate in human and ruminant metabolism. J. Nutr. 2005, 135, 1619–1625. [Google Scholar] [CrossRef]
- Sun, X.Q.; Fu, X.B.; Zhang, R.; Lu, Y.; Deng, Q.; Jiang, X.G.; Sheng, Z.Y. Relationship between plasma D (-)-lactate and intestinal damage after severe injuries in rats. World J. Gastroenterol. 2001, 7, 555–558. [Google Scholar] [CrossRef]
- Sobhian, B.; Kröpfl, A.; Hölzenbein, T.; Khadem, A.; Redl, H.; Bahrami, S. Increased circulating D-lactate levels predict risk of mortality after hemorrhage and surgical trauma in baboons. Shock 2012, 37, 473–477. [Google Scholar] [CrossRef] [PubMed]
- March, D.S.; Jones, A.W.; Thatcher, R.; Davison, G. The effect of bovine colostrum supplementation on intestinal injury and circulating intestinal bacterial DNA following exercise in the heat. Eur. J. Nutr. 2019, 58, 1441–1451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Païssé, S.; Valle, C.; Servant, F.; Courtney, M.; Burcelin, R.; Amar, J.; Lelouvier, B. Comprehensive description of blood microbiome from healthy donors assessed by 16 S targeted metagenomic sequencing. Transfusion 2016, 56, 1138–1147. [Google Scholar] [CrossRef] [PubMed]
- Nieman, D.; Kay, C.; Rathore, A.; Grace, M.; Strauch, R.; Stephan, E.; Sakaguchi, C.; Lila, M. Increased Plasma Levels of Gut-Derived Phenolics Linked to Walking and Running Following Two Weeks of Flavonoid Supplementation. Nutrients 2018, 10, 1718. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pals, K.L.; Chang, R.T.; Ryan, A.J.; Gisolfi, C.V. Effect of running intensity on intestinal permeability. J. Appl. Physiol. 1997, 82, 571–576. [Google Scholar] [CrossRef] [Green Version]
- JanssenDuijghuijsen, L.M.; Keijer, J.; Mensink, M.; Lenaerts, K.; Ridder, L.; Nierkens, S.; Kartaram, S.W.; Verschuren, M.C.; Pieters, R.H.; Bas, R.; et al. Adaptation of exercise-induced stress in well-trained healthy young men. Exp. Physiol. 2017, 102, 86–99. [Google Scholar] [CrossRef]
- Nieuwenhoven, V. The effect of physical exercise on parameters of gastrointestinal function. Neurogastroenterol. Motil. 1999, 11, 431–439. [Google Scholar] [CrossRef]
- Van Nieuwenhoven, M.A.; Brummer, R.J.; Brouns, F.J.P.H. Gastrointestinal function during exercise: Comparison of water, sports drink, and sports drink with caffeine. J. Appl. Physiol. 2000, 89, 1079–1085. [Google Scholar] [CrossRef] [Green Version]
- Van Nieuwenhoven, M.A.; Brouns, F.; Brummer, R.J.M. Gastrointestinal profile of symptomatic athletes at rest and during physical exercise. Eur. J. Appl. Physiol. 2004, 91, 429–434. [Google Scholar] [CrossRef]
- Marchbank, T.; Davison, G.; Oakes, J.R.; Ghatei, M.A.; Patterson, M.; Moyer, M.P.; Playford, R.J. The nutriceutical bovine colostrum truncates the increase in gut permeability caused by heavy exercise in athletes. Am. J. Physiol. Gastrointest. Liver Physiol. 2010, 300, G477–G484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davison, G.; Marchbank, T.; March, D.S.; Thatcher, R.; Playford, R.J. Zinc carnosine works with bovine colostrum in truncating heavy exercise-induced increase in gut permeability in healthy volunteers. Am. J. Clin. Nutr. 2016, 104, 526–536. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Snipe, R.M.; Khoo, A.; Kitic, C.M.; Gibson, P.R.; Costa, R.J. The impact of exertional-heat stress on gastrointestinal integrity, gastrointestinal symptoms, systemic endotoxin and cytokine profile. Eur. J. Appl. Physiol. 2018, 118, 389–400. [Google Scholar] [CrossRef] [PubMed]
- Zuhl, M.; Dokladny, K.; Mermier, C.; Schneider, S.; Salgado, R.; Moseley, P. The effects of acute oral glutamine supplementation on exercise-induced gastrointestinal permeability and heat shock protein expression in peripheral blood mononuclear cells. Cell Stress Chaperones 2015, 20, 85–93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Nieuwenhoven, M.A.; Vriens, B.E.P.J.; Brummer, R.J.; Brouns, F.J.P.H. Effect of dehydration on gastrointestinal function at rest and during exercise in humans. Eur. J. Appl. Physiol. 2000, 83, 578–584. [Google Scholar] [CrossRef] [PubMed]
- Lambert, G.P.; Lang, J.; Bull, A.; Pfeifer, P.C.; Eckerson, J.; Moore, G.; Lanspa, S.; O’Brien, J. Fluid restriction during running increases GI permeability. Int. J. Sports Med. 2008, 29, 194–198. [Google Scholar] [CrossRef] [PubMed]
- Ryan, A.J.; Chang, R.T.; Gisolfi, C.V. Gastrointestinal permeability following aspirin intake and prolonged running. Med. Sci. Sports Exerc. 1996, 28, 698–705. [Google Scholar] [CrossRef]
- Lambert, G.P.; Boylan, M.; Laventure, J.P.; Bull, A.; Lanspa, S. Effect of aspirin and ibuprofen on GI permeability during exercise. Int. J. Sports Med. 2007, 28, 722–726. [Google Scholar] [CrossRef]
- Lambert, G.P.; Broussard, L.J.; Mason, B.L.; Mauermann, W.J.; Gisolfi, C.V. Gastrointestinal permeability during exercise: Effects of aspirin and energy-containing beverages. J. Appl. Physiol. 2001, 90, 2075–2080. [Google Scholar] [CrossRef]
- Smetanka, R.D.; Lambert, C.P.; Murray, R.; Eddy, D.; Horn, M.; Gisolfi, C.V. Intestinal permeability in runners in the 1996 Chicago marathon. Int. J. Sport Nutr. Exerc. Metab. 1999, 9, 426–433. [Google Scholar] [CrossRef]
- Van Wijck, K.; Lenaerts, K.; Van Bijnen, A.A.; Boonen, B.; Van Loon, L.J.; Dejong, C.H.; Buurman, W.A. Aggravation of exercise-induced intestinal injury by Ibuprofen in athletes. Med. Sci. Sports Exerc. 2012, 44, 2257–2262. [Google Scholar] [CrossRef] [PubMed]
- Snipe, R.M.; Khoo, A.; Kitic, C.M.; Gibson, P.R.; Costa, R.J. The impact of mild heat stress during prolonged running on gastrointestinal integrity, gastrointestinal symptoms, systemic endotoxin and cytokine profiles. Int. J. Sports Med. 2018, 39, 255–263. [Google Scholar] [CrossRef] [PubMed]
- Shing, C.M.; Peake, J.M.; Lim, C.L.; Briskey, D.; Walsh, N.P.; Fortes, M.B.; Ahuja, K.D.; Vitetta, L. Effects of probiotics supplementation on gastrointestinal permeability, inflammation and exercise performance in the heat. Eur. J. Appl. Physiol. 2014, 114, 93–103. [Google Scholar] [CrossRef] [PubMed]
- Van Wijck, K.; Wijnands, K.A.; Meesters, D.M.; Boonen, B.; Van Loon, L.J.; Buurman, W.A.; Dejong, C.H.; Lenaerts, K.; Poeze, M. L-citrulline improves splanchnic perfusion and reduces gut injury during exercise. Med. Sci. Sports Exerc. 2014, 46, 2039–2046. [Google Scholar] [CrossRef] [PubMed]
- Buchman, A.L.; O’Brien, W.; Ou, C.N.; Rognerud, C.; Alvarez, M.; Dennis, K.; Ahn, C. The effect of arginine or glycine supplementation on gastrointestinal function, muscle injury, serum amino acid concentrations and performance during a marathon run. Int. J. Sports Med. 1999, 20, 315–321. [Google Scholar] [CrossRef]
- Snipe, R.M.; Costa, R.J. Does biological sex impact intestinal epithelial injury, small intestine permeability, gastrointestinal symptoms and systemic cytokine profile in response to exertional-heat stress? J. Sports Sci. 2018, 36, 2827–2835. [Google Scholar] [CrossRef]
- Snipe, R.M.; Khoo, A.; Kitic, C.M.; Gibson, P.R.; Costa, R.J. Carbohydrate and protein intake during exertional heat stress ameliorates intestinal epithelial injury and small intestine permeability. Appl. Physiol. Nutr. Metab. 2017, 42, 1283–1292. [Google Scholar] [CrossRef]
- March, D.S.; Marchbank, T.; Playford, R.J.; Jones, A.W.; Thatcher, R.; Davison, G. Intestinal fatty acid-binding protein and gut permeability responses to exercise. Eur. J. Appl. Physiol. 2017, 117, 931–941. [Google Scholar] [CrossRef] [Green Version]
- Zuhl, M.N.; Lanphere, K.R.; Kravitz, L.; Mermier, C.M.; Schneider, S.; Dokladny, K.; Moseley, P.L. Effects of oral glutamine supplementation on exercise–induced gastrointestinal permeability and tight junction protein expression. J. Appl. Physiol. 2013, 116, 183–191. [Google Scholar] [CrossRef] [Green Version]
- Buchman, A.L.; Killip, D.; Ou, C.N.; Rognerud, C.L.; Pownall, H.; Dennis, K.; Dunn, J.K. Short-term vitamin E supplementation before marathon running: A placebo-controlled trial. Nutrition 1999, 15, 278–283. [Google Scholar] [CrossRef]
- Pugh, J.N.; Sage, S.; Hutson, M.; Doran, D.A.; Fleming, S.C.; Highton, J.; Morton, J.P.; Close, G.L. Glutamine supplementation reduces markers of intestinal permeability during running in the heat in a dose–dependent manner. Eur. J. Appl. Physiol. 2017, 117, 2569–2577. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pugh, J.N.; Sparks, A.S.; Doran, D.A.; Fleming, S.C.; Langan-Evans, C.; Kirk, B.; Fearn, R.; Morton, J.P.; Close, G.L. Four weeks of probiotic supplementation reduces GI symptoms during a marathon race. Eur. J. Appl. Physiol. 2019, 119, 1491–1501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lambert, G.P.; Murray, R.; Eddy, D.; Scott, W.; Laird, R.; Gisolfi, C.V. Intestinal Permeability following the 1998 ironman triathlon. Med. Sci. Sports Exerc. 1999, 31, S318. [Google Scholar] [CrossRef]
- JanssenDuijghuijsen, L.; Van Norren, K.; Grefte, S.; Koppelman, S.; Lenaerts, K.; Keijer, J.; Witkamp, R.; Wichers, H. Endurance exercise increases intestinal uptake of the peanut allergen Ara h 6 after peanut consumption in humans. Nutrients 2017, 9, 84. [Google Scholar] [CrossRef] [PubMed]
- Lee, B.J.; Thake, C.D. Heat and Hypoxic Acclimation Increase Monocyte Heat Shock Protein 72 but Do Not Attenuate Inflammation following Hypoxic Exercise. Front. Physiol. 2017, 8, 811. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kartaram, S.; Mensink, M.; Teunis, M.; Schoen, E.; Witte, G.; Duijghuijsen, L.J.; Verschuren, M.; Mohrmann, K.; M’Rabet, L.; Knipping, K.; et al. Plasma citrulline concentration, a marker for intestinal functionality, reflects exercise intensity in healthy young men. Clin. Nutr. 2019, 38, 2251–2258. [Google Scholar] [CrossRef]
- Trommelen, J.; Fuchs, C.; Beelen, M.; Lenaerts, K.; Jeukendrup, A.; Cermak, N.; Van Loon, L. Fructose and sucrose intake increase exogenous carbohydrate oxidation during exercise. Nutrients 2017, 9, 167. [Google Scholar] [CrossRef] [Green Version]
- Jonvik, K.L.; Lenaerts, K.; Smeets, J.S.; Kolkman, J.J.; Van Loon, L.J.; Verdijk, L.B. Sucrose but Not Nitrate Ingestion Reduces Strenuous Cycling-induced Intestinal Injury. Med. Sci. Sports Exerc. 2019, 51, 436–444. [Google Scholar] [CrossRef] [Green Version]
- Karhu, E.; Forsgård, R.A.; Alanko, L.; Alfthan, H.; Pussinen, P.; Hämäläinen, E.; Korpela, R. Exercise and gastrointestinal symptoms: Running-induced changes in intestinal permeability and markers of gastrointestinal function in asymptomatic and symptomatic runners. Eur. J. Appl. Physiol. 2017, 117, 2519–2526. [Google Scholar] [CrossRef] [Green Version]
- Osborne, J.O.; Stewart, I.B.; Beagley, K.W.; Minett, G.M. The effect of cycling in the heat on gastrointestinal-induced damage and neuromuscular fatigue. Eur. J. Appl. Physiol. 2019, 119, 1829–1840. [Google Scholar] [CrossRef]
- Hill, G.W.; Gillum, T.L.; Lee, B.J.; Romano, P.A.; Schall, Z.J.; Hamilton, A.M.; Kuennen, M.R. Prolonged treadmill running in normobaric hypoxia causes gastrointestinal barrier permeability and elevates circulating levels of pro-and anti-inflammatory cytokines. Appl. Physiol. Nutr. Metab. 2019. [Google Scholar] [CrossRef] [PubMed]
- Costa, R.J.; Camoes-Costa, V.; Snipe, R.M.; Dixon, D.; Russo, I.; Huschtscha, Z. Impact of exercise-induced hypohydration on gastrointestinal integrity, function, symptoms, and systemic endotoxin and inflammatory profile. J. Appl. Physiol. 2019, 126, 1281–1291. [Google Scholar] [CrossRef] [PubMed]
- Sheahen, B.L.; Fell, J.W.; Zadow, E.K.; Hartley, T.F.; Kitic, C.M. Intestinal damage following short-duration exercise at the same relative intensity is similar in temperate and hot environments. Appl. Physiol. Nutr. Metab. 2018, 43, 1314–1320. [Google Scholar] [CrossRef] [PubMed]
- Snipe, R.M.; Costa, R.J. Does the temperature of water ingested during exertional-heat stress influence gastrointestinal injury, symptoms, and systemic inflammatory profile? J. Sci. Med Sport 2018, 21, 771–776. [Google Scholar] [CrossRef]
- Yeh, Y.J.; Law, L.Y.L.; Lim, C.L. Gastrointestinal response and endotoxemia during intense exercise in hot and cool environments. Eur. J. Appl. Physiol. 2013, 113, 1575–1583. [Google Scholar] [CrossRef]
- Edinburgh, R.M.; Hengist, A.; Smith, H.A.; Travers, R.L.; Koumanov, F.; Betts, J.A.; Thompson, D.; Walhin, J.P.; Wallis, G.A.; Hamilton, D.L.; et al. Preexercise breakfast ingestion versus extended overnight fasting increases postprandial glucose flux after exercise in healthy men. Am. J. Physiol. Endocrinol. Metab. 2018, 315, E1062–E1074. [Google Scholar] [CrossRef] [Green Version]
- Salvador, A.F.; McKenna, C.F.; Alamilla, R.A.; Cloud, R.M.; Keeble, A.R.; Miltko, A.; Scaroni, S.E.; Beals, J.W.; Ulanov, A.V.; Dilger, R.N.; et al. Potato ingestion is as effective as carbohydrate gels to support prolonged cycling performance. J. Appl. Physiol. 2019, 127, 1651–1659. [Google Scholar] [CrossRef] [Green Version]
- Nava, R.B.C.; Zuhl, M.N.; Moriarty, T.A.; Amorim, F.T.; Bourbeau, K.C.; Welch, A.M.; McCormick, J.J.; King, K.E.; Mermier, C.M. The effect of acute glutamine supplementation on markers of inflammation and fatigue during consecutive days of simulated wildland firefighting. J. Occup. Environ. Med. 2019, 61, e33–e42. [Google Scholar] [CrossRef]
- Lis, D.; Stellingwerff, T.; Kitic, C.K.; Ahuja, K.D.; Fell, J. No effects of a short-term gluten-free diet on performance in nonceliac athletes. Med. Sci. Sports Exerc. 2015, 47, 2563–2570. [Google Scholar] [CrossRef]
- Morrison, S.A.; Cheung, S.S.; Cotter, J.D. Bovine colostrum, training status, and gastrointestinal permeability during exercise in the heat: A placebo-controlled double-blind study. Appl. Physiol. Nutr. Metab. 2014, 39, 1070–1082. [Google Scholar] [CrossRef]
- Kashima, H.; Harada, N.; Miyamoto, K.; Fujimoto, M.; Fujita, C.; Endo, M.Y.; Kobayashi, T.; Miura, A.; Fukuba, Y. Timing of postexercise carbohydrate-protein supplementation: Roles of gastrointestinal blood flow and mucosal cell damage on gastric emptying in humans. J. Appl. Physiol. 2017, 123, 606–613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Osborne, J.O.; Stewart, I.B.; Beagley, K.W.; Borg, D.N.; Minett, G.M. Acute glutamine supplementation does not improve 20–km self–paced cycling performance in the heat. European J. Appl. Physiol. 2019, 119, 2567–2578. [Google Scholar] [CrossRef] [PubMed]
- Buckley, J.; Butler, R.; Southcott, E.; Brinkworth, G. Bovine colostrum supplementation during running training increases intestinal permeability. Nutrients 2009, 1, 224–234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szymanski, M.C.; Gillum, T.L.; Gould, L.M.; Morin, D.S.; Kuennen, M.R. Short-term dietary curcumin supplementation reduces gastrointestinal barrier damage and physiological strain responses during exertional heat stress. J. Appl. Physiol. 2017, 124, 330–340. [Google Scholar] [CrossRef] [PubMed]
- Raftery, T.; Martineau, A.R.; Greiller, C.L.; Ghosh, S.; McNamara, D.; Bennett, K.; Meddings, J.; O’Sullivan, M. Effects of vitamin D supplementation on intestinal permeability, cathelicidin and disease markers in Crohn’s disease: Results from a randomised double-blind placebo-controlled study. Un. Eur. Gastroenterol. J. 2015, 3, 294–302. [Google Scholar] [CrossRef] [PubMed]
- Stuempfle, K.J.; Valentino, T.; Hew–Butler, T.; Hecht, F.M.; Hoffman, M.D. Nausea is associated with endotoxemia during a 161-km ultramarathon. J. Sports Sci. 2016, 34, 1662–1668. [Google Scholar] [CrossRef] [PubMed]
- Bosenberg, A.T.; Brock-Utne, J.G.; Gaffin, S.L.; Wells, M.T.; Blake, G.T. Strenuous exercise causes systemic endotoxemia. J. Appl. Physiol. 1988, 65, 106–108. [Google Scholar] [CrossRef]
- Brock-Utne, J.G.; Gaffin, S.L.; Wells, M.T.; Gathiram, P.; Sohar, E.; James, M.F.; Morrell, D.F.; Norman, R.J. Endotoxaemia in exhausted runners after a long-distance race. South Afr. Med. J. Suid Afrik. Mediese Tydskr 1988, 73, 533–536. [Google Scholar]
- Moore, G.E.; Holbein, M.E.; Knochel, J.P. Exercise-associated collapse in cyclists is unrelated to endotoxemia. Med. Sci. Sports Exerc. 1995, 27, 1238–1242. [Google Scholar] [CrossRef]
- Kuennen, M.; Gillum, T.; Dokladny, K.; Bedrick, E.; Schneider, S.; Moseley, P. Thermotolerance and heat acclimation may share a common mechanism in humans. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2011, 301, R524–R533. [Google Scholar] [CrossRef] [Green Version]
- Jeukendrup, A.E.; Vet-Joop, K.; Sturk, A.; Stegen, J.H.J.C.; Senden, J.; Saris, W.H.M.; Wagenmakers, A.J.M. Relationship between gastro-intestinal complaints and endotoxaemia, cytokine release and the acute-phase reaction during and after a long-distance triathlon in highly trained men. Clin. Sc. 2000, 98, 47–55. [Google Scholar] [CrossRef] [Green Version]
- Gill, S.K.; Hankey, J.; Wright, A.; Marczak, S.; Hemming, K.; Allerton, D.M.; Ansley–Robson, P.; Costa, R.J.S. The impact of a 24-h ultra-marathon on circulatory endotoxin and cytokine profile. Int. J. Sports Med. 2015, 36, 688–695. [Google Scholar] [CrossRef] [PubMed]
- Selkirk, G.A.; McLellan, T.M.; Wright, H.E.; Rhind, S.G. Mild endotoxemia, NF-κB translocation, and cytokine increase during exertional heat stress in trained and untrained individuals. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2008, 295, R611–R623. [Google Scholar] [CrossRef] [PubMed]
- Gaskell, S.K.; Taylor, B.; Muir, J.; Costa, R.J. Impact of 24-h high and low fermentable oligo-di-mono-saccharide polyol diets on markers of exercise-induced gastrointestinal syndrome in response to exertional-heat stress. Appl. Physiol. Nutr. Metab. 2019. [Google Scholar] [CrossRef]
- Gill, S.K.; Allerton, D.M.; Ansley-Robson, P.; Hemmings, K.; Cox, M.; Costa, R.J. Does short-term high dose probiotic supplementation containing lactobacillus casei attenuate exertional-heat stress induced endotoxaemia and cytokinaemia? Int. J. Sport Nutr. Exerc. Metab. 2016, 26, 268–275. [Google Scholar] [CrossRef]
- Lim, C.L.; Pyne, D.; Horn, P.; Kalz, A.; Saunders, P.; Peake, J.; Suzuki, K.; Wilson, G.; Mackinnon, L.T. The effects of increased endurance training load on biomarkers of heat intolerance during intense exercise in the heat. Appl. Physiol. Nutr. Metab. 2009, 34, 616–624. [Google Scholar] [CrossRef]
- Antunes, B.M.; Campos, E.Z.; dos Santos, R.V.T.; Rosa–Neto, J.C.; Franchini, E.; Bishop, N.C.; Lira, F.S. Anti-inflammatory response to acute exercise is related with intensity and physical fitness. J. Cell. Biochem. 2019, 120, 5333–5342. [Google Scholar] [CrossRef] [Green Version]
- Guy, J.H.; Edwards, A.M.; Miller, C.M.; Deakin, G.B.; Pyne, D.B. Short-term reliability of inflammatory mediators and response to exercise in the heat. J. Sports Sci. 2017, 35, 1622–1628. [Google Scholar] [CrossRef]
- Ashton, T.; Young, I.S.; Davison, G.W.; Rowlands, C.C.; McEneny, J.; Van Blerk, C.; Jones, E.; Peters, J.R.; Jackson, S.K. Exercise-induced endotoxemia: The effect of ascorbic acid supplementation. Free Radical Biol. Med. 2003, 35, 284–291. [Google Scholar] [CrossRef]
- Guy, J.H.; Pyne, D.B.; Deakin, G.B.; Miller, C.M.; Edwards, A.M. Acclimation training improves endurance cycling performance in the heat without inducing endotoxemia. Front. Physiol. 2016, 7, 318. [Google Scholar] [CrossRef]
- Moss, J.N.; Bayne, F.M.; Castelli, F.; Naughton, M.R.; Reeve, T.C.; Trangmar, S.J.; Mackenzie, R.W.; Tyler, C.J. Short-term isothermic heat acclimation elicits beneficial adaptations but medium-term elicits a more complete adaptation. Eur. J. Appl. Physiol. 2020, 120, 243–254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Machado, P.; Caris, A.; Santos, S.; Silva, E.; Oyama, L.; Tufik, S.; Santos, R. Moderate exercise increases endotoxin concentration in hypoxia but not in normoxia: A controlled clinical trial. Medicine 2017, 96, e5504. [Google Scholar] [CrossRef] [PubMed]
- Moncada-Jiménez, J.; Plaisance, E.P.; Mestek, M.L.; Ratcliff, L.; Araya-Ramírez, F.; Taylor, J.K.; Grandjean, P.W.; AragónVargas, L.F. Duathlon performance unaltered by short-term changes in dietary fat and carbohydrates. Int. J. Sport Nutr. Exerc. Metab. 2009, 19, 47–60. [Google Scholar] [CrossRef] [PubMed]
- Dokladny, K.; Zuhl, M.N.; Moseley, P.L. Intestinal epithelial barrier function and tight junction proteins with heat and exercise. J. Appl. Physiol. 2015, 120, 692–701. [Google Scholar] [CrossRef]
- Grootjans, J.; Lenaerts, K.; Buurman, W.A.; Dejong, C.H.; Derikx, J.P. Life and death at the mucosal-luminal interface: New perspectives on human intestinal ischemia-reperfusion. World J. Gastroenterol. 2016, 22, 2760. [Google Scholar] [CrossRef]
- De Punder, K.; Pruimboom, L. Stress induces endotoxemia and low-grade inflammation by increasing barrier permeability. Front. Immunol. 2015, 6, 223. [Google Scholar] [CrossRef] [Green Version]
- Sawka, M.N.; Leon, L.R.; Montain, S.J.; Sonna, L.A. Integrated physiological mechanisms of exercise performance, adaptation, and maladaptation to heat stress. Compr. Physiol. 2011, 1, 1883–1928. [Google Scholar]
- Byrne, C.; Lim, C.L. The ingestible telemetric body core temperature sensor: A review of validity and exercise applications. Br. J. Sports Med. 2007, 41, 126–133. [Google Scholar] [CrossRef]
- Dokladny, K.; Moseley, P.L.; Ma, T.Y. Physiologically relevant increase in temperature causes an increase in intestinal epithelial tight junction permeability. Am. J. Physiol. Gastrointest. Liver Physiol. 2006, 290, G204–G212. [Google Scholar] [CrossRef]
- Oliver, S.R.; Phillips, N.A.; Novosad, V.L.; Bakos, M.P.; Talbert, E.E.; Clanton, T.L. Hyperthermia induces injury to the intestinal mucosa in the mouse: Evidence for an oxidative stress mechanism. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2012, 302, R845–R853. [Google Scholar] [CrossRef] [Green Version]
- Ikari, A.; Nakano, M.; Suketa, Y.; Harada, H.; Takagi, K. Reorganization of ZO-1 by sodium-dependent glucose transporter activation after heat stress in LLC-PK1 cells. J. Cell. Physiol. 2005, 203, 471–478. [Google Scholar] [CrossRef] [PubMed]
- Yang, P.C.; He, S.H.; Zheng, P.Y. Investigation into the signal transduction pathway via which heat stress impairs intestinal epithelial barrier function. J. Gastroenterol. Hepatol. 2007, 22, 1823–1831. [Google Scholar] [CrossRef] [PubMed]
- Ter Steege, R.W.F.; Kolkman, J.J. The pathophysiology and management of gastrointestinal symptoms during physical exercise, and the role of splanchnic blood flow. Aliment. Pharmacol. Ther. 2012, 35, 516–528. [Google Scholar] [CrossRef] [PubMed]
- Ahlborg, G.U.N.V.O.R.; Weitzberg, E.; Lundberg, J. Metabolic and vascular effects of circulating endothelin-1 during moderately heavy prolonged exercise. J. Appl. Physiol. 1995, 78, 2294–2300. [Google Scholar] [CrossRef] [PubMed]
- Kenney, W.L.; Ho, C.W. Age alters regional distribution of blood flow during moderate-intensity exercise. J. Appl. Physiol. 1995, 79, 1112–1119. [Google Scholar] [CrossRef] [PubMed]
- Otte, J.A.; Oostveen, E.; Geelkerken, R.H.; Groeneveld, A.J.; Kolkman, J.J. Exercise induces gastric ischemia in healthy volunteers: A tonometry study. J. Appl. Physiol. 2001, 91, 866–871. [Google Scholar] [CrossRef]
- Friedman, E.S.; Bittinger, K.; Esipova, T.V.; Hou, L.; Chau, L.; Jiang, J.; Mesaros, C.; Lund, P.J.; Liang, X.; FitzGerald, G.A.; et al. Microbes vs.chemistry in the origin of the anaerobic gut lumen. Proc. Natl. Acad. Sci. USA 2018, 115, 4170–4175. [Google Scholar] [CrossRef] [Green Version]
- JanssenDuijghuijsen, L.M.; Grefte, S.; de Boer, V.C.; Zeper, L.; van Dartel, D.A.; van der Stelt, I.; Bekkenkamp-Grovenstein, M.; van Norren, K.; Wichers, H.J.; Keijer, J. Mitochondrial ATP depletion disrupts Caco-2 monolayer integrity and internalizes claudin 7. Front. Physiol. 2017, 8, 794. [Google Scholar] [CrossRef] [Green Version]
- Turner, M.D.; Nedjai, B.; Hurst, T.; Pennington, D.J. Cytokines and chemokines: At the crossroads of cell signalling and inflammatory disease. Biochim. Biophys. Acta (BBA) Mol. Cell Res. 2014, 1843, 2563–2582. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, K. Cytokine response to exercise and its modulation. Antioxidants 2018, 7, 17. [Google Scholar] [CrossRef] [Green Version]
- Al-Sadi, R.; Guo, S.; Dokladny, K.; Smith, M.A.; Ye, D.; Kaza, A.; Watterson, D.M.; Ma, T.Y. Mechanism of interleukin-1β induced-increase in mouse intestinal permeability in vivo. J. Interferon Cytokine Res. 2012, 32, 474–484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Sadi, R.; Guo, S.; Ye, D.; Ma, T.Y. TNF-α modulation of intestinal epithelial tight junction barrier is regulated by ERK1/2 activation of Elk-1. Am. J. Pathol. 2013, 183, 1871–1884. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Sadi, R.; Ye, D.; Boivin, M.; Guo, S.; Hashimi, M.; Ereifej, L.; Ma, T.Y. Interleukin-6 modulation of intestinal epithelial tight junction permeability is mediated by JNK pathway activation of claudin-2 gene. PLoS ONE 2014, 9, e85345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fortes, M.B.; Di, U.F.; Dolci, A.; Junglee, N.A.; Crockford, M.J.; West, L.; Hillier-Smith, R.; Macdonald, J.H.; Walsh, N.P. Muscle-damaging exercise increases heat strain during subsequent exercise heat stress. Med. Sci. Sports Exerc. 2013, 45, 1915–1924. [Google Scholar] [CrossRef]
- Dolci, A.; Fortes, M.B.; Walker, F.S.; Haq, A.; Riddle, T.; Walsh, N.P. Repeated muscle damage blunts the increase in heat strain during subsequent exercise heat stress. Eur. J. Appl. Physiol. 2015, 115, 1577–1588. [Google Scholar] [CrossRef] [Green Version]
- Cavaillon, J.M.; Annane, D. Invited review: Compartmentalization of the inflammatory response in sepsis and SIRS. J. Endotoxin Res. 2006, 12, 151–170. [Google Scholar] [CrossRef]
- Ghanim, H.; Sia, C.L.; Upadhyay, M.; Korzeniewski, K.; Viswanathan, P.; Abuaysheh, S.; Mohanty, P.; Dandona, P. Orange juice neutralizes the proinflammatory effect of a high-fat, high-carbohydrate meal and prevents endotoxin increase and Toll-like receptor expression. Am. J. Clin. Nutr. 2010, 91, 940–949. [Google Scholar] [CrossRef] [Green Version]
- Deopurkar, R.; Ghanim, H.; Friedman, J.; Abuaysheh, S.; Sia, C.L.; Mohanty, P.; Viswanathan, P.; Chaudhuri, A.; Dandona, P. Differential effects of cream, glucose, and orange juice on inflammation, endotoxin, and the expression of Toll-like receptor-4 and suppressor of cytokine signaling-3. Diabetes Care 2010, 33, 991–997. [Google Scholar] [CrossRef] [Green Version]
- Ghanim, H.; Abuaysheh, S.; Sia, C.L.; Korzeniewski, K.; Chaudhuri, A.; Fernandez–Real, J.M.; Dandona, P. 2009. Increase in plasma endotoxin concentrations and the expression of Toll-like receptors and suppressor of cytokine signaling-3 in mononuclear cells after a high-fat, high-carbohydrate meal: Implications for insulin resistance. Diabetes Care 2009, 32, 2281–2287. [Google Scholar] [CrossRef] [Green Version]
- Deniz, T.; Agalar, C.; Ozdogan, M.; Comu, F.; Emirdogan, M.; Taskin, S.; Saygun, O.; Agalar, F. Oral carbohydrate solution ameliorates endotoxemia–induced splanchnic ischemia. Dig. Dis. Sci. 2007, 52, 287–291. [Google Scholar] [CrossRef]
- Ramadass, B.; Dokladny, K.; Moseley, P.L.; Patel, Y.R.; Lin, H.C. Sucrose co-administration reduces the toxic effect of lectin on gut permeability and intestinal bacterial colonization. Dig. Dis. Sci. 2010, 55, 2778–2784. [Google Scholar] [CrossRef] [PubMed]
- David, L.A.; Maurice, C.F.; Carmody, R.N.; Gootenberg, D.B.; Button, J.E.; Wolfe, B.E.; Ling, A.V.; Devlin, A.S.; Varma, Y.; Fischbach, M.A.; et al. Diet rapidly and reproducibly alters the human gut microbiome. Nature 2014, 505, 559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gentilcore, D.; Nair, N.S.; Vanis, L.; Rayner, C.K.; Meyer, J.H.; Hausken, T.; Horowitz, M.; Jones, K.L. Comparative effects of oral and intraduodenal glucose on blood pressure, heart rate, and splanchnic blood flow in healthy older subjects. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2009, 297, R716–R722. [Google Scholar] [CrossRef] [PubMed]
- Lerner, A.; Matthias, T. Changes in intestinal tight junction permeability associated with industrial food additives explain the rising incidence of autoimmune disease. Autoimmun. Rev. 2015, 14, 479–489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Erridge, C.; Attina, T.; Spickett, C.M.; Webb, D.J. A high-fat meal induces low-grade endotoxemia: Evidence of a novel mechanism of postprandial inflammation. Am. J. Clin. Nutr. 2007, 86, 1286–1292. [Google Scholar] [CrossRef]
- Pöchmüller, M.; Schwingshackl, L.; Colombani, P.C.; Hoffmann, G. A systematic review and meta-analysis of carbohydrate benefits associated with randomized controlled competition-based performance trials. J. Int. Soc. Sports Nutr. 2016, 13, 27. [Google Scholar] [CrossRef] [Green Version]
- McCartney, D.; Desbrow, B.; Irwin, C. Post-exercise ingestion of carbohydrate, protein and water: A systematic review and meta-analysis for effects on subsequent athletic performance. Sports Med. 2018, 48, 379–408. [Google Scholar] [CrossRef]
- Bermon, S.; Castell, L.M.; Calder, P.C.; Bishop, N.C.; Blomstrand, E.; Mooren, F.C.; Krüger, K.; Kavazis, A.N.; Quindry, J.C.; Senchina, D.S.; et al. Consensus Statement Immunonutrition and Exercise. Exerc. Immunol. Rev. 2017, 23, 8–50. [Google Scholar]
- Costa, R.J.; Miall, A.; Khoo, A.; Rauch, C.; Snipe, R.; Camões–Costa, V.; Gibson, P. Gut-training: The impact of two weeks repetitive gut-challenge during exercise on gastrointestinal status, glucose availability, fuel kinetics, and running performance. Appl. Physiol. Nutr. Metab. 2017, 42, 547–557. [Google Scholar] [CrossRef] [Green Version]
- Rehrer, N.J.; Goes, E.; DuGardeyn, C.; Reynaert, H.; DeMeirleir, K. Effect of carbohydrate on portal vein blood flow during exercise. Int. J. Sports Med. 2005, 26, 171–176. [Google Scholar] [CrossRef]
- Sessions, J.; Bourbeau, K.; Rosinski, M.; Szczygiel, T.; Nelson, R.; Sharma, N.; Zuhl, M. Carbohydrate gel ingestion during running in the heat on markers of gastrointestinal distress. Eur. J. Sport Sci. 2016, 16, 1064–1072. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.H.; Kim, H. The roles of glutamine in the intestine and its implication in intestinal diseases. Int. J. Mol. Sci. 2017, 18, 1051. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Y.P.; Jiang, Z.M.; Sun, Y.H.; Wang, X.R.; Ma, E.L.; Wilmore, D. The effect of supplemental enteral glutamine on plasma levels, gut function, and outcome in severe burns: A randomized, double-blind, controlled clinical trial. J. Parenter. Enteral Nutr. 2003, 27, 241–245. [Google Scholar] [CrossRef] [PubMed]
- Peng, X.; Yan, H.; You, Z.; Wang, P.; Wang, S. Effects of enteral supplementation with glutamine granules on intestinal mucosal barrier function in severe burned patients. Burns 2004, 30, 135–139. [Google Scholar] [CrossRef]
- Zhou, Q.; Verne, M.L.; Fields, J.Z.; Lefante, J.J.; Basra, S.; Salameh, H.; Verne, G.N. Randomised placebo–controlled trial of dietary glutamine supplements for postinfectious irritable bowel syndrome. Gut 2019, 68, 996–1002. [Google Scholar]
- Shu, X.L.; Yu, T.T.; Kang, K.; Zhao, J. Effects of glutamine on markers of intestinal inflammatory response and mucosal permeability in abdominal surgery patients: A meta-analysis. Exp. Ther. Med. 2016, 12, 3499–3506. [Google Scholar] [CrossRef] [Green Version]
- Akobeng, A.K.; Miller, V.; Stanton, J.; Elbadri, A.M.; Thomas, A.G. Double-blind randomized controlled trial of glutamine-enriched polymeric diet in the treatment of active Crohn’s disease. J. Pediatr. Gastroenterol. Nutr. 2000, 30, 78–84. [Google Scholar] [CrossRef]
- Benjamin, J.; Makharia, G.; Ahuja, V.; Rajan, K.A.; Kalaivani, M.; Gupta, S.K.D.; Joshi, Y.K. Glutamine and whey protein improve intestinal permeability and morphology in patients with Crohn’s disease: A randomized controlled trial. Dig. Dis. Sci. 2012, 57, 1000–1012. [Google Scholar] [CrossRef]
- Rhoads, J.M.; Argenzio, R.A.; Chen, W.U.N.I.A.N.; Rippe, R.A.; Westwick, J.K.; Cox, A.D.; Berschneider, H.M.; Brenner, D.A. L-glutamine stimulates intestinal cell proliferation and activates mitogen-activated protein kinases. Am. J. Physiol. Gastrointest. Liver Physiol. 1997, 272, G943–G953. [Google Scholar] [CrossRef]
- Singleton, K.D.; Wischmeyer, P.E. Oral glutamine enhances heat shock protein expression and improves survival following hyperthermia. Shock 2006, 25, 295–299. [Google Scholar] [CrossRef]
- Xue, H.; Sufit, A.J.; Wischmeyer, P.E. Glutamine therapy improves outcome of in vitro and in vivo experimental colitis models. J. Parenter. Enteral Nutr. 2011, 35, 188–197. [Google Scholar] [CrossRef] [PubMed]
- Dai, Z.L.; Li, X.L.; Xi, P.B.; Zhang, J.; Wu, G.; Zhu, W.Y. L-Glutamine regulates amino acid utilization by intestinal bacteria. Amino Acids 2013, 45, 501–512. [Google Scholar] [CrossRef] [PubMed]
- Harward, T.R.; Coe, D.; Souba, W.W.; Klingman, N.; Seeger, J.M. Glutamine preserves gut glutathione levels during intestinal ischemia/reperfusion. J. Surg. Res. 1994, 56, 351–355. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Lewis, P.; Samuelson, D.; Liboni, K.; Neu, J. Glutamine regulates Caco-2 cell tight junction proteins. Am. J. Physiol. Gastrointest. Liver Physiol. 2004, 287, G726–G733. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beutheu, S.; Ghouzali, I.; Galas, L.; Déchelotte, P.; Coëffier, M. Glutamine and arginine improve permeability and tight junction protein expression in methotrexate-treated Caco-2 cells. Clin. Nutr. 2013, 32, 863–869. [Google Scholar] [CrossRef] [PubMed]
- Ahmadi, A.R.; Rayyani, E.; Bahreini, M.; Mansoori, A. The effect of glutamine supplementation on athletic performance, body composition, and immune function: A systematic review and a meta-analysis of clinical trials. Clin. Nutr. 2019, 38, 1076–1091. [Google Scholar] [CrossRef]
- Gleeson, M. Dosing and efficacy of glutamine supplementation in human exercise and sport training. J. Nutr. 2008, 138, 2045S–2049S. [Google Scholar] [CrossRef]
- Rathe, M.; Müller, K.; Sangild, P.T.; Husby, S. Clinical applications of bovine colostrum therapy: A systematic review. Nutr. Rev. 2014, 72, 237–254. [Google Scholar] [CrossRef]
- Uruakpa, F.O.; Ismond, M.A.H.; Akobundu, E.N.T. Colostrum and its benefits: A review. Nutr. Res. 2002, 22, 755–767. [Google Scholar] [CrossRef]
- Prosser, C.; Stelwagen, K.; Cummins, R.; Guerin, P.; Gill, N.; Milne, C. Reduction in heat-induced gastrointestinal hyperpermeability in rats by bovine colostrum and goat milk powders. J. Appl. Physiol. 2004, 96, 650–654. [Google Scholar] [CrossRef] [Green Version]
- Playford, R.J.; Floyd, D.N.; Macdonald, C.E.; Calnan, D.P.; Adenekan, R.O.; Johnson, W.; Goodlad, R.A.; Marchbank, T. Bovine colostrum is a health food supplement which prevents NSAID induced gut damage. Gut 1999, 44, 653–658. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eslamian, G.; Ardehali, S.H.; Baghestani, A.R.; Shariatpanahi, Z.V. Effects of early enteral bovine colostrum supplementation on intestinal permeability in critically ill patients: A randomized, double-blind, placebo-controlled study. Nutrition 2019, 60, 106–111. [Google Scholar] [CrossRef] [PubMed]
- Bodammer, P.; Kerkhoff, C.; Maletzki, C.; Lamprecht, G. Bovine colostrum increases pore-forming claudin-2 protein expression but paradoxically not ion permeability possibly by a change of the intestinal cytokine milieu. PLoS ONE 2013, 8, e64210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, Z.; Macdonald, C.; Wicks, A.C.; Holt, M.P.; Floyd, D.; Ghosh, S.; Wright, N.A.; Playford, R.J. Use of the ‘nutriceutical’, bovine colostrum, for the treatment of distal colitis: Results from an initial study. Aliment. Pharmacol. Ther. 2002, 16, 1917–1922. [Google Scholar] [CrossRef]
- Antonio, J.; Sanders, M.S.; Van Gammeren, D. The effects of bovine colostrum supplementation on body composition and exercise performance in active men and women. Nutrition 2001, 17, 243–247. [Google Scholar] [CrossRef]
- Jones, A.W.; March, D.S.; Curtis, F.; Bridle, C. Bovine colostrum supplementation and upper respiratory symptoms during exercise training: A systematic review and meta-analysis of randomised controlled trials. BMC Sports Sci. Med. Rehabil. 2016, 8, 21. [Google Scholar] [CrossRef] [Green Version]
- Shing, C.M.; Hunter, D.C.; Stevenson, L.M. Bovine colostrum supplementation and exercise performance. Sports Med. 2009, 39, 1033–1054. [Google Scholar] [CrossRef]
- McKenna, Z.; Berkemeier, Q.; Naylor, A.; Kleint, A.; Gorini, F.; Ng, J.; Kim, J.K.; Sullivan, S.; Gillum, T. Bovine colostrum supplementation does not affect plasma I-FABP concentrations following exercise in a hot and humid environment. Eur. J. Appl. Physiol. 2017, 117, 2561–2567. [Google Scholar] [CrossRef]
- Hałasa, M.; Maciejewska, D.; Baśkiewicz–Hałasa, M.; Machaliński, B.; Safranow, K.; Stachowska, E. Oral supplementation with bovine colostrum decreases intestinal permeability and stool concentrations of zonulin in athletes. Nutrients 2017, 9, 370. [Google Scholar] [CrossRef]
- Jasion, V.S.; Burnett, B.P. Survival and digestibility of orally-administered immunoglobulin preparations containing IgG through the gastrointestinal tract in humans. Nutr. J. 2015, 14, 22. [Google Scholar] [CrossRef] [Green Version]
- Davison, G.; Jones, A.W.; Marchbank, T.; Playford, R.J. Oral bovine colostrum supplementation does not increase circulating insulin-like growth factor-1 concentration in healthy adults: Results from short-and long-term administration studies. Eur. J. Nutr. 2019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lundberg, J.O.; Carlström, M.; Weitzberg, E. Metabolic effects of dietary nitrate in health and disease. Cell Metab. 2018, 28, 9–22. [Google Scholar] [CrossRef] [PubMed]
- Castillo, L.; DeRojas, T.C.; Chapman, T.E.; Vogt, J.; Burke, J.L.F.; Tannenbaum, S.R.; Young, V.R. Splanchnic metabolism of dietary arginine in relation to nitric oxide synthesis in normal adult man. Proc. Natl. Acad. Sci. USA 1993, 90, 193–197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petersson, J.; Phillipson, M.; Jansson, E.A.; Patzak, A.; Lundberg, J.O.; Holm, L. Dietary nitrate increases gastric mucosal blood flow and mucosal defense. Am. J. Physiol. Gastrointest. Liver Physiol. 2007, 292, G718–G724. [Google Scholar] [CrossRef]
- Tanaka, A.; Araki, H.; Komoike, Y.; Hase, S.; Takeuchi, K. Inhibition of both COX-1 and COX-2 is required for development of gastric damage in response to nonsteroidal antiinflammatory drugs. J. Physiol. Paris 2001, 95, 21–27. [Google Scholar] [CrossRef]
- Batista, M.A.; Nicoli, J.R.; dos Santos Martins, F.; Nogueira Machado, J.A.; Esteves Arantes, R.M.; Pacífico Quirino, I.E.; Davisson Correia, M.I.T.; Cardoso, V.N. Pretreatment with citrulline improves gut barrier after intestinal obstruction in mice. J. Parenter. Enteral Nutr. 2012, 36, 69–76. [Google Scholar] [CrossRef]
- Sukhotnik, I.; Helou, H.; Mogilner, J.; Lurie, M.; Bernsteyn, A.; Coran, A.G.; Shiloni, E. Oral arginine improves intestinal recovery following ischemia-reperfusion injury in rat. Pediatr. Surg. Int. 2005, 21, 191–196. [Google Scholar] [CrossRef] [Green Version]
- Gou, L.; Zhang, L.; Yin, C.; Jia, G.; Yin, X.; Zhuang, X.; Xu, X.; Liu, Y. Protective effect of l-citrulline against acute gastric mucosal lesions induced by ischemia-reperfusion in rats. Can. J. Physiol. Pharmacol. 2011, 89, 317–327. [Google Scholar] [CrossRef]
- Rubanyi, G.M.; Ho, E.H.; Cantor, E.H.; Lumma, W.C.; Botelho, L.H.P. Cytoprotective function of nitric oxide: Inactivation of superoxide radicals produced by human leukocytes. Biochem. Biophys. Res. Commun. 1991, 181, 1392–1397. [Google Scholar] [CrossRef]
- Kubes, P.; Suzuki, M.; Granger, D.N. Nitric oxide: An endogenous modulator of leukocyte adhesion. Proc. Natl. Acad. Sci. USA 1991, 88, 4651–4655. [Google Scholar] [CrossRef] [Green Version]
- Beutheu, S.; Ouelaa, W.; Guérin, C.; Belmonte, L.; Aziz, M.; Tennoune, N.; Bôle-Feysot, C.; Galas, L.; Déchelotte, P.; Coëffier, M. Glutamine supplementation, but not combined glutamine and arginine supplementation, improves gut barrier function during chemotherapy-induced intestinal mucositis in rats. Clin. Nutr. 2014, 33, 694–701. [Google Scholar] [CrossRef] [PubMed]
- Jones, A.M.; Thompson, C.; Wylie, L.J.; Vanhatalo, A. Dietary nitrate and physical performance. Ann. Rev. Nutr. 2018, 38, 303–328. [Google Scholar] [CrossRef] [PubMed]
- Bailey, S.J.; Blackwell, J.R.; Lord, T.; Vanhatalo, A.; Winyard, P.G.; Jones, A.M. L-citrulline supplementation improves O2 uptake kinetics and high-intensity exercise performance in humans. J. Appl. Physiol. 2015, 119, 385–395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Costa, K.A.; Soares, A.D.N.; Wanner, S.P.; Santos, R.D.G.C.D.; Fernandes, S.O.A.; Martins, F.D.S.; Nicoli, J.R.; Coimbra, C.C.; Cardoso, V.N. L-arginine supplementation prevents increases in intestinal permeability and bacterial translocation in male Swiss mice subjected to physical exercise under environmental heat stress. J. Nutr. 2013, 144, 218–223. [Google Scholar] [CrossRef] [Green Version]
- McMahon, N.F.; Leveritt, M.D.; Pavey, T.G. The effect of dietary nitrate supplementation on endurance exercise performance in healthy adults: A systematic review and meta-analysis. Sports Med. 2017, 47, 735–756. [Google Scholar] [CrossRef] [Green Version]
- Kuennen, M.; Jansen, L.; Gillum, T.; Granados, J.; Castillo, W.; Nabiyar, A.; Christmas, K. Dietary nitrate reduces the O 2 cost of desert marching but elevates the rise in core temperature. Eur. J. Appl. Physiol. 2015, 115, 2557–2569. [Google Scholar] [CrossRef]
- McQuillan, J.A.; Casadio, J.R.; Dulson, D.K.; Laursen, P.B.; Kilding, A.E. The effect of nitrate supplementation on cycling performance in the heat in well-trained cyclists. Int. J. Sports Physiol. Perform. 2018, 13, 50–56. [Google Scholar] [CrossRef]
- Parvez, S.; Malik, K.A.; Ah Kang, S.; Kim, H.Y. Probiotics and their fermented food products are beneficial for health. J. Appl. Microbiol. 2006, 100, 1171–1185. [Google Scholar] [CrossRef]
- van Hemert, S.; Verwer, J.; Schütz, B. Clinical studies evaluating effects of probiotics on parameters of intestinal barrier function. Adv. Microbiol. 2013, 3, 212. [Google Scholar] [CrossRef] [Green Version]
- Bron, P.A.; Van Baarlen, P.; Kleerebezem, M. Emerging molecular insights into the interaction between probiotics and the host intestinal mucosa. Nat. Rev. Microbiol. 2012, 10, 66. [Google Scholar] [CrossRef]
- Resta–Lenert, S.; Barrett, K.E. Probiotics and commensals reverse TNF-α-and IFN-γ-induced dysfunction in human intestinal epithelial cells. Gastroenterology 2006, 130, 731–746. [Google Scholar] [CrossRef]
- Hsieh, C.Y.; Osaka, T.; Moriyama, E.; Date, Y.; Kikuchi, J.; Tsuneda, S. Strengthening of the intestinal epithelial tight junction by Bifidobacterium bifidum. Physiol. Rep. 2015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- West, N.P.; Pyne, D.B.; Peake, J.M.; Cripps, A.W. Probiotics, immunity and exercise: A review. Exerc. Immunol. Rev. 2009, 15, e26. [Google Scholar]
- Pyne, D.B.; West, N.P.; Cox, A.J.; Cripps, A.W. Probiotics supplementation for athletes-clinical and physiological effects. Eur. J. Sport Sci. 2015, 15, 63–72. [Google Scholar] [CrossRef]
- Suzuki, T.; Hara, H. Quercetin enhances intestinal barrier function through the assembly of zonnula occludens-2, occludin, and claudin-1 and the expression of claudin-4 in Caco-2 cells. J. Nutr. 2009, 139, 965–974. [Google Scholar] [CrossRef]
- Roberts, J.; Suckling, C.; Peedle, G.; Murphy, J.; Dawkins, T.; Roberts, M. An exploratory investigation of endotoxin levels in novice long distance triathletes, and the effects of a multi-strain probiotic/prebiotic, antioxidant intervention. Nutrients 2016, 8, 733. [Google Scholar] [CrossRef] [Green Version]
- Carbuhn, A.; Reynolds, S.; Campbell, C.; Bradford, L.; Deckert, J.; Kreutzer, A.; Fry, A. Effects of probiotic (Bifidobacterium longum 35624) supplementation on exercise performance, immune modulation, and cognitive outlook in Division I female swimmers. Sports 2018, 6, 116. [Google Scholar] [CrossRef] [Green Version]
- Axelrod, C.L.; Brennan, C.J.; Cresci, G.; Paul, D.; Hull, M.; Fealy, C.E.; Kirwan, J.P. UCC118 supplementation reduces exercise-induced gastrointestinal permeability and remodels the gut microbiome in healthy humans. Physiol. Rep. 2019. [Google Scholar] [CrossRef] [Green Version]
- Karl, J.P.; Margolis, L.M.; Madslien, E.H.; Murphy, N.E.; Castellani, J.W.; Gundersen, Y.; Hoke, A.V.; Levangie, M.W.; Kumar, R.; Chakraborty, N.; et al. Changes in intestinal microbiota composition and metabolism coincide with increased intestinal permeability in young adults under prolonged physiological stress. Am. J. Physiol. Gastrointest. Liver Physiol. 2017, 312, G559–G571. [Google Scholar] [CrossRef] [Green Version]
- De Vries, J.H.; Hollman, P.C.; Meyboom, S.; Buysman, M.N.; Zock, P.L.; van Staveren, W.A.; Katan, M.B. Plasma concentrations and urinary excretion of the antioxidant flavonols quercetin and kaempferol as biomarkers for dietary intake. Am. J. Clin. Nutr. 1998, 68, 60–65. [Google Scholar] [CrossRef] [Green Version]
- Sukhotnik, I.; Moati, D.; Shaoul, R.; Loberman, B.; Pollak, Y.; Schwartz, B. Quercetin prevents small intestinal damage and enhances intestinal recovery during methotrexate-induced intestinal mucositis of rats. Food Nutr. Res. 2018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amasheh, M.; Schlichter, S.; Amasheh, S.; Mankertz, J.; Zeitz, M.; Fromm, M.; Schulzke, J.D. Quercetin enhances epithelial barrier function and increases claudin-4 expression in Caco-2 cells. J. Nutr. 2008, 138, 1067–1073. [Google Scholar] [CrossRef] [Green Version]
- Dokladny, K.; Ye, D.; Kennedy, J.C.; Moseley, P.L.; Ma, T.Y. Cellular and molecular mechanisms of heat stress-induced up-regulation of occludin protein expression: Regulatory role of heat shock factor-1. Am. J. Pathol. 2008, 172, 659–670. [Google Scholar] [CrossRef] [Green Version]
- Freedman, J.E.; Parker Iii, C.; Li, L.; Perlman, J.A.; Frei, B.; Ivanov, V.; Deak, L.R.; Iafrati, M.D.; Folts, J.D. Select flavonoids and whole juice from purple grapes inhibit platelet function and enhance nitric oxide release. Circulation 2001, 103, 2792–2798. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oteiza, P.I.; Fraga, C.G.; Mills, D.A.; Taft, D.H. Flavonoids and the gastrointestinal tract: Local and systemic effects. Mol. Asp. Med. 2018, 61, 41–49. [Google Scholar] [CrossRef] [PubMed]
- Valenzano, M.C.; DiGuilio, K.; Mercado, J.; Teter, M.; To, J.; Ferraro, B.; Mixson, B.; Manley, I.; Baker, V.; Moore, B.A.; et al. Remodeling of tight junctions and enhancement of barrier integrity of the CACO-2 intestinal epithelial cell layer by micronutrients. PLoS ONE 2015, 10, e0133926. [Google Scholar] [CrossRef] [Green Version]
- Hosokawa, N.; Hirayoshi, K.; Kudo, H.; Takechi, H.; Aoike, A.; Kawai, K.; Nagata, K. Inhibition of the activation of heat shock factor in vivo and in vitro by flavonoids. Mol. Cell. Biol. 1992, 12, 3490–3498. [Google Scholar] [CrossRef] [Green Version]
- Dokladny, K.; Wharton, W.; Lobb, R.; Ma, T.Y.; Moseley, P.L. Induction of physiological thermotolerance in MDCK monolayers: Contribution of heat shock protein 70. Cell Stress Chaperones 2006, 11, 268. [Google Scholar] [CrossRef]
- Sergent, T.; Piront, N.; Meurice, J.; Toussaint, O.; Schneider, Y.J. Anti-inflammatory effects of dietary phenolic compounds in an in vitro model of inflamed human intestinal epithelium. Chem. Biol. Interact. 2010, 188, 659–667. [Google Scholar] [CrossRef]
- Knapik, J.J.; Austin, K.G.; Farina, E.K.; Lieberman, H.R. Dietary supplement use in a large, representative sample of the US armed forces. J. Acad. Nutr. Dietetics 2018, 118, 1370–1388. [Google Scholar] [CrossRef]
- Myburgh, K.H. Polyphenol supplementation: Benefits for exercise performance or oxidative stress? Sports Med. 2014, 44, 57–70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nieman, D.C.; Mitmesser, S.H. Potential impact of nutrition on immune system recovery from heavy exertion: A metabolomics perspective. Nutrients 2017, 9, 513. [Google Scholar] [CrossRef] [Green Version]
- Somerville, V.; Bringans, C.; Braakhuis, A. Polyphenols and performance: A systematic review and meta-analysis. Sports Med. 2017, 47, 1589–1599. [Google Scholar] [CrossRef]
- Takei, M. Development of polaprezinc research. Yakugaku Zasshi J. Pharm. Soc. Jpn. 2012, 132, 271–277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsukura, T.; Tanaka, H. Applicability of Zinc Complex of L-Carnosine for Medical Use. Biochemistry 2000, 65, 817–823. [Google Scholar]
- Watari, I.; Oka, S.; Tanaka, S.; Aoyama, T.; Imagawa, H.; Shishido, T.; Yoshida, S.; Chayama, K. Effectiveness of polaprezinc for low-dose aspirin-induced small-bowel mucosal injuries as evaluated by capsule endoscopy: A pilot randomized controlled study. BMC Gastroenterol. 2013, 13, 108. [Google Scholar] [CrossRef] [Green Version]
- Roohani, N.; Hurrell, R.; Kelishadi, R.; Schulin, R. Zinc and its importance for human health: An integrative review. J. Res. Med. Sci. 2013, 18, 144. [Google Scholar]
- Sale, C.; Artioli, G.G.; Gualano, B.; Saunders, B.; Hobson, R.M.; Harris, R.B.C. Carnosine: From exercise performance to health. Amino Acids 2013, 44, 1477–1491. [Google Scholar] [CrossRef]
- Mahmood, A.; Fitzgerald, A.J.; Marchbank, T.; Ntatsaki, E.; Murray, D.; Ghosh, S.; Playford, R.J. Zinc carnosine, a health food supplement that stabilises small bowel integrity and stimulates gut repair processes. Gut 2007, 56, 168–175. [Google Scholar] [CrossRef] [Green Version]
- Omatsu, T.; Naito, Y.; Handa, O.; Mizushima, K.; Hayashi, N.; Qin, Y.; Harusato, A.; Hirata, I.; Kishimoto, E.; Okada, H.; et al. Reactive oxygen species-quenching and anti-apoptotic effect of polaprezinc on indomethacin-induced small intestinal epithelial cell injury. J. Gastroenterol. 2010, 45, 692–702. [Google Scholar] [CrossRef]
- Fujii, Y.; Matsura, T.; Kai, M.; Kawasaki, H.; Yamada, K. Protection by polaprezinc, an anti-ulcer drug, against indomethacin-induced apoptosis in rat gastric mucosal cells. Jpn. J. Pharmacol. 2000, 84, 63–70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, H.S.; Lim, J.Y.; Chun, H.J.; Lee, M.; Kim, E.S.; Keum, B.; Seo, Y.S.; Jeen, Y.T.; Um, S.H.; Lee, H.S.; et al. The effect of polaprezinc on gastric mucosal protection in rats with ethanol-induced gastric mucosal damage: Comparison study with rebamipide. Life Sci. 2013, 93, 69–77. [Google Scholar] [CrossRef]
- Mayorga, E.J.; Kvidera, S.K.; Horst, E.A.; Al-Qaisi, M.; Dickson, M.J.; Seibert, J.T.; Lei, S.; Keating, A.F.; Ross, J.W.; Rhoads, R.P.; et al. Effects of zinc amino acid complex on biomarkers of gut integrity and metabolism during and following heat stress or feed restriction in pigs. J. Anim. Sci. 2018, 96, 4173–4185. [Google Scholar] [CrossRef]
- Pearce, S.C.; Sanz Fernandez, M.V.; Torrison, J.; Wilson, M.E.; Baumgard, L.H.; Gabler, N.K. Dietary organic zinc attenuates heat stress–induced changes in pig intestinal integrity and metabolism. J. Anim. Sci. 2015, 93, 4702–4713. [Google Scholar] [CrossRef]
- Shah, M.; Zaneb, H.; Masood, S.; Khan, R.U.; Mobashar, M.; Khan, I.; Din, S.; Khan, M.S.; Rehman, H.U.; Tinelli, A. Single or Combined Applications of Zinc and Multi–strain Probiotic on Intestinal Histomorphology of Broilers Under Cyclic Heat Stress. Probiotics Antimicrob. Proteins 2019. [Google Scholar] [CrossRef]
- Weng, X.; Monteiro, A.P.A.; Guo, J.; Li, C.; Orellana, R.M.; Marins, T.N.; Bernard, J.K.; Tomlinson, D.J.; DeFrain, J.M.; Wohlgemuth, S.E.; et al. Effects of heat stress and dietary zinc source on performance and mammary epithelial integrity of lactating dairy cows. J. Dairy Sci. 2018, 101, 2617–2630. [Google Scholar] [CrossRef] [Green Version]
- Saunders, B.; Elliott–Sale, K.; Artioli, G.G.; Swinton, P.A.; Dolan, E.; Roschel, H.; Sale, C.; Gualano, B. β-alanine supplementation to improve exercise capacity and performance: A systematic review and meta-analysis. Br. J. Sports Med. 2017, 51, 658–669. [Google Scholar] [CrossRef]
- Sakae, K.; Yanagisawa, H. Oral treatment of pressure ulcers with polaprezinc (zinc L-carnosine complex): 8-week open-label trial. Biol. Trace Element Res. 2014, 158, 280–288. [Google Scholar] [CrossRef]
- Barbalho, S.M.; Goulart, A.R.; Quesada, K.; Bechara, M.D. Inflammatory bowel disease: Can omega-3 fatty acids really help? Ann. Gastroenterol. 2016, 29, 37–43. [Google Scholar]
- Carlson, J.; Slavin, J. Health benefits of fibre, prebiotics and probiotics: A review of intestinal health and related health claims. Qual. Assur. Safety Crops Foods 2016, 8, 539–554. [Google Scholar] [CrossRef]


{kind=link}
| Technique | Sample | Method | Site | Limitations |
|---|---|---|---|---|
| Active Techniques | ||||
| Dual-Sugar Absorption Test (DSAT) | Urine or blood | HPLC (+) MS | Small GI Integrity | Gold-standard. High reliability. Time-consuming (5 h urine, >2.5 h blood). No standard protocol with exercise. Well studied. |
| Multi-Sugar Absorption Test (MSAT) | Urine or blood | HLPC (+) MS | Entire GI Integrity | Gold-Standard. Segmental GI integrity. Time-consuming (5 h urine, >2.5 h blood). No standard protocol with exercise. Few studies. |
| Polyethylene Glycol (PEG) Absorption Test | Urine | HLPC (+) MS | Entire GI Integrity | Validated against MSAT. Can include multiple weight PEGs (e.g., 100, 400, 1000, 4000 kDa). Time-consuming (5 h urine). Few studies. |
| Passive Techniques | ||||
| Intestinal Fatty Acid Binding Protein (I-FABP) | Urine or Blood | ELISA | Epithelial injury | Tissue specific (duodenum and jejunum). Short half-life (11 min). Weak correlations with DSAT. Well studied. |
| Ileal Bile-Acid Binding Protein (I-BABP) | Urine or Blood | ELISA | Epithelial injury | Tissue specific (ileum). Few studies. Weak correlations with I-FABP. Few studies. |
| Diamine Oxidase (DAO), α-Glutathione s-Transferase (α-GST), Smooth Muscle 22 (SM22) | Blood | ELISA | Epithelial injury | Non-tissue specific. Few studies. |
| Claudin-3 (CLDN3) | Urine or Blood | ELISA | TJ Integrity | Non-tissue specific. Few studies. |
| Zonulin | Blood or Faeces | ELISA | TJ Integrity | Non-tissue specific. Assay cross-reactivity (complement C3). Moderate studies. |
| Endotoxin (LPS) | Blood | LAL assay | MT | Tissue specific. Sample contamination causes false-positives. Hepatic removal and receptor binding cause false-negatives. Well studied. |
| LPS Binding Protein (LBP) | Blood | ELISA | MT | Tissue specific. Lower risk of false-positives than endotoxin. Indirect marker of endotoxin exposure. Influenced by hepatic production. Long half-life (12–14 h). Few studies. |
| Soluable-CD14 (sCD14-ST) | Blood | ELISA | MT | Tissue specific. Lower risk of false positives than endotoxin. Influenced by hepatic production and monocytes shedding. Few studies. |
| D-lactate | Blood | ELISA | MT | Predominately tissue specific. Potentially influenced by methylglyoxal metabolism. Few studies. |
| 16s Bacterial rDNA (bactDNA) | Blood | Real-time PCR assay | MT | Tissue specific. Novel. Lower risk of false-positives than endotoxin. Few studies. |
| Author | Subjects | Exercise Protocol | Peak TCore (°C) | Mean HR (bpm) | Biofluid, DSAT L/R or L/M (Timepoint) |
|---|---|---|---|---|---|
| van Nieuwenhoven et al. [110] | 10 male (MT) | 90 min cycling at 70% Wattmax (fasted) in Tamb 19 °C (RH = N/A) | N/A | N/A | Urine L/R (5 h): 0.007 s |
| van Nieuwenhoven et al. [117] | 10 male (MT) | 90 min cycling at 70% Wattmax (fasted) in Tamb 19 °C (RH = N/A) | 38.8 | N/A | Urine L/R (5 h): 0.008 nb,c |
| Nieman et al. [107] | 20 male and female (UT) | 45 min walking uphill (5% grade) at 60% VO2max (fasted) in Tamb not reported | N/A | 132 | Urine L/R (5 h): 0.009 nb,c |
| Smetanka et al. [122] | 8 male (HT) | Chicago marathon (42.2 km) in Tamb (fed) 22 °C (48% RH) | N/A | N/A | Urine L/R (5 h): 0.020 ns |
| Shing et al. [125] | 10 male (HT) | ~33 min running to fatigue at 80% VE (fed) in Tamb 35 °C (40% RH) | 39.4 | 172 | Urine L/R (5 h): 0.022 nb,c |
| Janssen-Duijghuijsen et al. [109] | 11 male (HT) | 90 min cycling at 50% wattmax (fed) in Tamb not reported following a sleep-low glycogen depletion regime | N/A | N/A | Urine L/R (5 h): ~0.022 ns Plasma L/R (1 h): ~0.110 s |
| Snipe et al. [115,124] | 6 male and 4 female (MT) | 120 min running at 60% VO2max (fed) in Tamb 22 °C (44% RH) | 38.5 | ~150 | Urine L/R (5 h): 0.025 nb |
| Snipe et al. [124] | 6 male and 4 female (MT) | 120 min running at 60% VO2max (fed) in Tamb 30 °C (25% RH) | 38.6 | ~155 | Urine L/R (5 h): 0.026 nb |
| van Wijck et al. [126] | 10 male (MT) | 60 min cycling at 70% wattmax (fasted) in Tamb not reported | N/A | N/A | Urine L/R (2 h): 0.027 nb,c |
| Buchman et al. [127] | 17 male and 2 female | Competitive Marathon (fed) in Tamb 2 °C with freezing rain | N/A | N/A | Urine L/R (6 h): 0.030 ns,c |
| Ryan et al. [119] | 7 males (MT) | 60 min running at 68% VO2max (fasted) in Tamb not reported | N/A | N/A | Urine L/M (6 h): 0.029 ns |
| van Nieuwenh-oven et al. [112] | 9 male and 1 female (MT) | 90 min cycling at 70% Wattmax (fasted) in Tamb 19 °C (RH = N/A) | N/A | N/A | Urine L/R (5 h): 0.030 ns |
| van Wijck et al. [123] | 9 male (MT) | 60 min cycling at 70% wattmax (fasted) in Tamb not reported | N/A | N/A | Urine L/R (2 h): 0.030 s,c |
| Pugh et al. [88] | 11 male (MT-HT) | 18 × 400 m sprint at 120% VO2max (fed) in Tamb not reported | N/A | N/A | Urine L/R (2h): 0.030 ns Serum L/R (2 h): ~0.051 s |
| Snipe and Costa [128] | 11 male (MT) | 120 min running at 60% VO2max (fed) in Tamb 35 °C (25% RH) | 39.1 | ~150 | Urine L/R (5 h): 0.030 nb |
| Snipe and Costa [128] | 13 female (MT) | 120 min running at 60% VO2max (fed) in Tamb 35 °C (25% RH) | 38.8 | ~155 | Urine L/R (5 h): 0.028 nb |
| Snipe et al. (Part B) [115] | 6 male and 4 female (MT) | 120 min running at 60% VO2max (fed) in Tamb 35 °C (26% RH) | 39.6 | ~170 | Urine L/R (5 h): 0.032 nb |
| Snipe et al. [129] | 6 male and 5 female (MT) | 120 min running at 60% VO2max (fed) in Tamb 35 °C (30% RH) | 39.3 | 159 | Urine L/R (5 h): 0.034 nb,c |
| March et al. [130] | 9 male (MT) | 20 min running at 80% VO2peak (fasted) in Tamb 22 °C (37% RH) | 38.4 | 170 | Urine L/R (5 h): 0.035 s,c |
| Pals et al. (Part A) [108] | 5 male and 1 female (MT) | 60 min running at 40% VO2peak (fasted) in Tamb 22 °C (50% RH) | 38.0 | N/A | Urine L/R (5 h): 0.036 ns |
| Marchbank et al. [113] | 12 male (MT) | 20 min running to fatigue at 80% VO2max (fasted) in Tamb not reported | 38.3 | N/A | Urine L/R (5 h): 0.038 s,c |
| van Nieuwenh-oven et al. [111] | 9 male and 1 female (MT) | 90 min running at 70% VO2max (fasted) in Tamb 19 °C (RH = N/A) | N/A | N/A | Urine L/R (5 h): 0.040 s |
| van Wijck et al. [86] | 6 male (HT) | 60 min cycling at 70% wattmax (fasted) in Tamb not reported | N/A | N/A | Urine L/R (5 h): 0.040 ns Plasma L/R (2.4 h): 0.060 s |
| Lambert et al. (Part A) [118] | 11 male and 9 female (MT) | 60 min running at 70% VO2max (fasted) in Tamb 22 °C (48% RH) | 38.5 | N/A | Urine L/R (5 h): 0.049 ns,c |
| Lambert et al. [121] | 13 male and 4 female (HT) | 60 min running at 70% VO2max (fasted) in Tamb 22 °C (48% RH) | 38.3 | N/A | Urine L/R (5 h): 0.050 nb,c |
| Zuhl et al. [131] | 4 male and 3 female (LT/MT) | 60 min running at 70% VO2max (fasted) in Tamb 30 °C (12–20% RH) | 39.4 | N/A | Urine L/R (5 h): 0.060 nb,c |
| Zuhl et al. [116] | 2 male and 5 female (LT/MT) | 60 min running at 70% VO2max (fasted) in Tamb 30 °C (12–20% RH) | 39.5 | N/A | Urine L/R (5 h): 0.060 nb,c |
| Lambert et al. (Part B) [118] | 11 male and 9 female (MT) | 60 min running at 70% VO2max (fasted) in Tamb 22 °C (48% RH) without fluid ingestion | 38.5 | N/A | Urine L/R (5 h): 0.063 s,c |
| Pals et al. (Part B) [108] | 5 male and 1 female (MT) | 60 min running at 40% VO2peak (fasted) in Tamb 22 °C (50% RH) | 38.7 | N/A | Urine L/R (5 h): 0.064 ns |
| Lambert et al. [120] | 8 male (MT) | 60 min running at 70% VO2max (fasted) in Tamb 22 °C (48% RH) | 38.3 | N/A | Urine L/R (5 h): 0.065 nb,c |
| Buchman et al. [132] | 15 male and female (LT-HT) | Road marathon (42.2 km) (fed) in Tamb not reported | N/A | N/A | Urine L/M (6 h): 0.070 ns,c |
| Pugh et al. [133] | 10 male (MT) | 60 min at 70% VO2max running (fasted) in Tamb 30 °C (4–45% RH) | 38.5 | 82.5% of max | Serum L/R (2 h): ~0.080 s,c |
| Pugh et al. [134] | 10 male and 2 female (MT) | 42.4 km track marathon (247 ± 47 min; fed) in Tamb 16–17 °C (N/A RH) | N/A | ~160 | Serum L/R (1 h) 0.081 (37%) s,c |
| Lambert et al. [135] | 12 female (LT-HT) | Hawaii Ironman (fed) in Tamb not reported | N/A | N/A | Urine L/R (5 h): 0.087 nb |
| Davison et al. [114] | 8 male (MT/HT) | 20 min running to fatigue at 80% VO2max (fasted) in Tamb not reported | 39.3 | ~170 | Urine L/R (5 h): 0.098 s,c |
| Janssen-Duijghuijsen et al. [136] | 4 male and 6 female (LT) | 60 min cycling at 70% wattmax (fed) in Tamb not reported | N/A | N/A | Plasma L/R (1 h): ~0.100 s |
| Lambert et al. [135] | 29 male (LT-HT) | Hawaii Ironman (fed) in Tamb not reported | N/A | N/A | Urine L/R (5 h): 0.105 nb |
| Pals et al. (Part C) [108] | 5 male and 1 female (MT) | 60 min running at 40% VO2peak (fasted) in Tamb 22 °C (50% RH) | 39.6 | N/A | Urine L/R (5 h): 0.107 s |
| Author | Subjects | Exercise Protocol | Peak TCore (°C) | Mean HR (bpm) | I-FABP (Δ Pre-to-Post Exercise) |
|---|---|---|---|---|---|
| Janssen-Duijghuijsen et al. [109] | 11 male (HT) | 90 min cycling at 50% wattmax (fed) in Tamb not reported following a “sleep-low” glycogen depletion regime | N/A | N/A | ~−90 pg·mL−1 (~−65%) c |
| Kartaram et al. (Part A) [138] | 15 male (MT) | 60 min cycling at 50% wattmax (fed) in Tamb not reported | N/A | N/A | ~-50 pg·mL−1 (~−10%) ns |
| Lee and Thake (Part A) [137] | 7 male (MT) | 60 min cycling at 50% VO2max (fed) in Tamb 18 °C (35% RH) on day one of temperate acclimation | 37.9 | 133 | 28 pg·mL−1 (8%) ns,c |
| Trommelen et al. [139] | 10 male (HT) | 180 min cycling at 50% wattmax (fasted) in Tamb 18–22 °C (55–65% RH) | N/A | N/A | N/A pg·mL−1 (20%) ns,c |
| Edinburgh et al. (Part A) [148] | 12 male (MT) | 60 min cycling at 50% VO2max (fed) in Tamb 18 °C (35% RH) | N/A | N/A | 70 pg·mL−1 (34%) s |
| Edinburgh et al. (Part B) [148] | 12 male (MT) | 60 min cycling at 50% VO2max (fasted) in Tamb 18 °C (35% RH) | N/A | N/A | 88 pg·mL−1 (20%) s |
| Osborne et al. (Part A) [142] | 8 male (MT-HT) | 30 min cycling at 50/70% Wattmax, then 30 min at 50% wattmax (fasted) in Tamb 20 °C (55% RH) | 38.5 | 139 | 138 pg·mL−1 (29%) ns |
| Salvador et al. 2019 [149] | 12 male (MT-HT) | 120 min cycling at 60% VO2max (fed) then 30–40 min (20 km) time trial in Tamb not reported | 37.9 | ~168 | N/A pg·mL−1 (~50%) s, c |
| van Wijck et al. [126] | 10 male (MT) | 60 min cycling at 70% wattmax (fasted) in Tamb not reported | N/A | N/A | 153 pg·mL−1 (72%) s |
| Nava et al. [150] | 7 male and 4 female (LT-MT) | 56 min mixed intensity (~55% VO2max) discontinuous firefighting exercises (fed) in Tamb 38 °C (35% RH) on day one of two | 38.7 | ~161 | ~160 pg·mL−1 (23%) ns, c |
| Van Wijck et al. [123] | 9 male (MT) | 60 min cycling at 70% wattmax (fasted) in Tamb not reported | N/A | N/A | 179 pg·mL−1 (61%) s |
| Lee et al. (Part C) [137] | 7 male (MT) | 60 min cycling at 50% VO2max (fed) in Tamb 18 °C (35% RH) and FiO2 = 0.14% on day one of hypoxic acclimation | 38.2 | 149 | 193 pg·mL−1 (43%) s,c |
| Lis et al. [151] | 13 male and female (MT) | 45 min cycling at 70% wattmax and 15 min cycling time trial (fed) in 20 °C (40% RH) | N/A | 168 | 210 pg·mL−1 (223%) s,c |
| Pugh et al. [134] | 10 male (MT) | 60 min at 70% VO2max running (fasted) in Tamb 30 °C (4–45% RH) | 38.5 | 82.5% of HR max | 250 pg·mL−1 (71%) s,c |
| Snipe et al. (Part A) [115,124] | 6 male and 4 female (MT) | 120 min running at 60% VO2max (fed) in Tamb 22 °C (44% RH) | 38.5 | ~150 | 274 pg·mL−1 (127%) s |
| Sheahen et al. (Part A) [145] | 12 male (MT) | 45 min running at 70% VO2max (fasted) in Tamb 20 °C (40% RH) | 38.2 | 165 | 281 pg·mL−1 (49%) s |
| Lee et al. (Part B) [137] | 7 male (MT) | 60 min cycling at 50% VO2max (fed) in Tamb 40 °C (25% RH) on day one of heat acclimation | 38.7 | 151 | 282 pg·mL−1 (76%) s,c |
| Morrison et al. (Part B) [152] | 8 male (UT) | 30 min cycling at 50% heart rate reserve (HRR), 30 min jogging at 80% HRR and 30 min running time trial (fed) in Tamb 30 °C (50% RH) | 38.6 | N/A | 283 pg·mL−1 (276%) s,c |
| Barberio et al. [72] | 9 male (MT) | ~24 min running at 78% VO2max (fed) in Tamb 40 °C (40% RH) prior to heat acclimation | 39.0 | N/A | 297 pg·mL−1 (46%) s,c |
| Hill et al. [143] | 10 male (MT) | 60 min running at 65% VO2max (fasted) in Tamb not reported | N/A | ~170 | 300 pg·mL−1 (50%) ns,c |
| van Wijck et al. [86] | 15 male (HT) | 60 min cycling at 70% wattmax (fasted) in Tamb not reported | N/A | N/A | 306 pg·mL−1 (61%) s |
| Kashima et al. [153] | 5 male and 3 female (MT) | 30 intermittent 20 s cycle sprints at 120% wattmax, with 40 s recovery between each (fed) in 23 °C (40% RH) | N/A | 150 | 343 pg·mL−1 (266%) s |
| Pugh et al. [88] | 11 male (MT-HT) | 18 × 400 m sprint at 120% VO2max (fed) in Tamb not reported | N/A | N/A | 348 pg·mL−1 (72%) s |
| March et al. [130] | 9 male (MT) | 20 min running at 80% VO2peak (fasted) in Tamb 22 °C (37% RH) | 38.4 | 170 | 350 pg·mL−1 (61%) s,c |
| Janssen-Duijghuijsen et al. [136] | 4 male and 6 female (LT) | 60 min cycling at 70% wattmax (fed) in Tamb not reported | N/A | N/A | ~350 pg·mL−1 (~77%) s,c |
| Sheahen et al. (Part B) [145] | 12 male (MT) | 45 min running at 70% VO2max (fasted) in Tamb 30 °C (40% RH) | 38.3 | 163 | 369 pg·mL−1 (63%) s |
| Costa et al. [144] | 11 male (MT-HT) | 120 min running at 70% VO2max (fed) in Tamb 25 °C (35% RH) | N/A | 148 | 371 pg·mL−1 (86%) ns,c |
| Osborne et al. [154] | 12 male (MT-HT) | 33 min (20 km) cycling time trial (fasted) in 35 °C (50% RH) | 39 | 167 | 441 pg·mL−1 (83%) s,c |
| Kartaram et al. (Part B) [138] | 15 male (MT) | 60 min cycling at 70% wattmax (fed) in Tamb not reported | N/A | N/A | ~500 pg·mL−1 (~66%) s |
| Kartaram et al. (Part C) [138] | 15 male (MT) | 60 min cycling at 85/55% wattmax (fed) in Tamb not reported | N/A | N/A | ~500 pg·mL−1 (~66%) s |
| McKenna et al. [155] | 10 male (MT) | 46 min running at 95% VE threshold (fasted) in Tamb 40 °C (50% RH) | 39.7 | N/A | 516 pg·mL−1 (52%) s,c |
| Karhu et al. [141] | 17 male (MT-HT) | 90 min running at 80% of best 10 km race time (fed) in Tamb not reported | N/A | N/A | 531 pg·mL−1 (151%) s |
| Snipe and Costa [146] | 6 male and 6 female (MT) | 120 min running at 60% VO2max (fed) in Tamb 30 °C (35% RH) | 38.8 | 160 | 573 pg·mL−1 (184%) s,c |
| Snipe et al. (Part B) [124] | 6 male and 4 female (MT) | 120 min running at 60% VO2max (fed) in Tamb 30 °C (25% RH) | 38.6 | ~155 | ~580 pg·mL−1 (184%) |
| Hill et al. [143] | 10 male (MT) | 60 min running at 65% VO2max (fasted) in Tamb not reported (FiO2 = 13.5%) | N/A | ~170 | 700 pg·mL−1 (168%) ns,c |
| Osborne et al. (Part B) [142] | 8 Male (MT-HT) | 30 min cycling at 50/70% Wattmax, then 30 min at 50% wattmax (fasted) in Tamb 35 °C (53% RH) | 39.5 | 159 | 608 pg·mL−1 (140%) s |
| Szymanski et al. [156] | 6 male and 2 female (LT/MT) | 60 min running at 68% VO2max (fasted) in Tamb 37 °C (25% RH) | 39.0 | 174 | 800 pg·mL−1 (87%) s,c |
| Morrison et al. (Part A) [152] | 7 male (HT) | 30 min cycling at 50% heart rate reserve (HRR), 30 min jogging at 80% HRR and 30 min running time trial (fed) in Tamb 30 °C (50% RH) | 38.6 | N/A | 806 pg·mL−1 (663%) s,c |
| Snipe et al. [129] | 6 male and 5 female (MT) | 120 min running at 60% VO2max (fed) in Tamb 35 °C (30% RH) | 39.3 | 159 | 897 pg·mL−1 (288%) s,c |
| Snipe et al. (Part B) [115] | 6 male and 4 female (MT) | 120 min running at 60% VO2max (fed) in Tamb 35 °C (26% RH) | 39.6 | ~170 | 1230 pg·mL−1 (432%) s |
| Pugh et al. [134] | 10 male and 2 female (MT) | 42.4 km track marathon (247 ± 47 min; fed) in Tamb 16-17 °C (N/A RH) | N/A | ~160 | 1246 pg·mL−1 (371%) s, c |
| March et al. [105] | 12 male (MT) | 60 min running at 70% VO2max (fasted) in Tamb 30 °C (60% RH) | 39.3 | 170 | 1263 pg·mL−1 (407%) s, c |
| Snipe and Costa [157] | 11 male (MT) | 120 min running at 60% VO2max (fed) in Tamb 35 °C (25% RH) | 39.1 | ~150 | 1389 pg·mL−1 (479%) s |
| Snipe et al. [158] | 13 female (MT) | 120 min running at 60% VO2max (fed) in Tamb 35 °C (25% RH) | 38.8 | ~155 | 1445 pg·mL−1 (479%) s |
| Jonvik et al. [140] | 16 male (HT) | 60 min cycling at 70% wattmax (fasted) in Tamb not reported | N/A | N/A | 1745 pg·mL−1 (249%) s |
| Gaskell et al. [134] | 10 male and 8 female (MT-HT) | 120 min running at 60% VO2max (fed) in Tamb 35 °C (25% RH) | 38.6 | ~151 | 1805 pg·mL−1 (710%) s, c |
| Author | Subjects | Exercise Protocol | Peak TCore (°C) | Mean HR (bpm) | Endotoxin (Δ Pre-to-Post Exercise) |
|---|---|---|---|---|---|
| Antunes et al. [169] | 19 male (MT) | 56 ± 7 min cycling at 90% of first ventilatory threshold (fasted) in 22.1 °C (55% RH) | N/A | 141 | −3 pg·mL−1 (−3%) ns |
| Yeh et al. (Part B) [147] | 15 male and 1 female (LT) | 60 min running at 70% VO2max (fed) in Tamb 22 °C (66% RH) | 38.4 | ~145 | −1.1 pg·mL−1 (−10%) ns |
| Zuhl et al. [116] | 2 male and 5 female (LT/MT) | 60 min running at 70% VO2max (fasted) in Tamb 30 °C (12–20% RH) | 39.5 | N/A | −0.2 pg·mL−1 (−7%) ns,c |
| Osborne et al. (Part A) [142] | 8 Male (MT-HT) | 30 min cycling at 50/70% Wattmax, then 30 min at 50% wattmax (fasted) in Tamb 20 °C (55% RH) | 38.5 | 165 | 0.1 pg·mL−1 (1%) ns,# |
| Osborne et al. (Part B) [142] | 8 Male (MT-HT) | 30 min cycling at 50/70% Wattmax, then 30 min at 50% wattmax (fasted) in Tamb 35 °C (53% RH) | 39.5 | 182 | 0.2 pg·mL−1 (1%) s,# |
| Karhu et al. [141] | 17 males (MT-HT) | 90 min running at 80% of best 10 km race time (fed) in Tamb not reported | N/A | N/A | 0.3 pg·mL−1 (~1%) ns,c |
| Kuennen et al. [162] | 8 male (MT) | 100 min walking (6.3 km·h−1) at 50% VO2max (fasted) in Tamb 46.5 °C (20% RH) | 39.3 | N/A | ~0.5 pg·mL−1 (10%) ns,c |
| Ng et al. [73] | 30 males (HT) | Half-marathon (fed) in Tamb 27 °C (84% RH) | 40.7 | 172 | 0.6 pg·mL−1 (32%) s |
| Jeukendrup et al. [163] | 29 male and 1 female (HT) | Ironman (3.8 km swim; 185 km cycle; 42.2 km run) (fed) in Tamb 9–32 °C | N/A | N/A | 1.7 pg·mL−1 (666%) s |
| Guy et al. [170] | 20 male (LT-MT) | 10 min cycling at 50%, 60%, and 70% wattmax, then 5 km (fasted) in Tamb 35 °C (70% RH) | 38.9 | 160 | 2 pg·mL−1 (9%) ns |
| Selkirk et al. (Part B) [125] | 12 male (HT) | To fatigue (~122 min) uphill walk at 4.5 km.h−1 (fasted) in Tamb 40 °C (30% RH) | 39.7 | 156 | ~3 pg·mL−1 (200%) s |
| Shing et al. [165] | 10 male (HT) | ~33 min running to fatigue at 80% VE (fed) in Tamb 35 °C (40% RH) | 39.4 | 172 | 4 pg·mL−1 (15%) s |
| Snipe et al. (Part A) [115,124] | 6 male and 4 female (MT) | 120 min running at 60% VO2max (fed) in Tamb 22 °C (44% RH) | 38.5 | ~150 | 4.1 pg·mL−1 (5%) ns |
| Yeh et al. (Part B) [147] | 15 male and 1 female (LT) | 60 min running at 70% VO2max (fed) in Tamb 33 °C (50% RH) | 39.3 | ~145 | 5 pg·mL−1 (54%) s |
| Antunes et al. (Part B) [169] | 19 male (MT) | 45 ± 18 min cycling at midpoint between first and second ventilatory threshold (fasted) in 22.1 °C (55% RH) | N/A | 162 | 5 pg·mL−1 (7%) ns |
| Antunes et al. (Part C) [169] | 19 male (MT) | 10 ± 9 min cycling at midpoint between second ventilatory threshold and maximal aerobic power (fasted) in 22.1 °C (55% RH) | N/A | 180 | 6 pg·mL−1 (5%) ns |
| Ashton et al. [171] | 10 males (LT) | VO2max test (~15 min) on cycle ergometer (fasted) in Tamb not reported | N/A | N/A | 9.4 pg·mL−1 (72%) s |
| Snipe et al. (Part B) [115] | 6 male and 4 female (MT) | 120 min running at 60% VO2max (fed) in Tamb 35 °C (26% RH) | 39.6 | ~170 | 9.8 pg·mL−1 (11%) s |
| Gill et al. [167] | 8 male (MT-HT) | 120 min running at 60% VO2max (fed) in Tamb 32 °C (34% RH) | 38.6 | 165 | 10 pg·mL−1 (4%) ns, c |
| Snipe et al. [129] | 6 male and 5 female (MT) | 120 min running at 60% VO2max (fed) in Tamb 35 °C (30% RH) | 39.3 | 159 | 10 pg·mL−1 (N/A%) nb |
| Selkirk et al. (Part A) [125] | 11 male (LT-MT) | To fatigue (~106 min) uphill walk at 4.5 km.h−1 (fasted) in Tamb 40 °C (30% RH) | 39.1 | 164 | ~10 pg·mL−1 (300%) s |
| Lim et al. (Part B) [168] | 9 male (HT) | To fatigue (time not given) at 70% VO2max (fed) in Tamb 35 °C (40% RH) | 39.5 | N/A | 13 pg·mL−1 (92%) s,c |
| Guy et al. [172] | 8 male (LT) | 10 min cycling at 50%, 60%, and 70% wattmax, then 5 km (fasted) in Tamb 35 °C (70% RH) | 38.6 | 161 | 16 pg·mL−1 (9%) ns,c,# |
| Gill et al. [71] | 13 male and 6 female (HT) | Multistage ultra-marathon stage 1 (37 km) (fed) in Tamb 32–40 °C (32–40% RH) | N/A | N/A | 40 pg·mL−1 (14%) s |
| Barberio et al. [72] | 9 male (MT) | ~24 min running at 78% VO2max (fed) in Tamb 40 °C (40% RH) prior to heat acclimation | 39.0 | N/A | 40 pg·mL−1 (57%) s,c |
| Moss et al. [173] | 9 male (HT) | 45 min cycling at 40% PPO (unstated prandial state) in Tamb 40 °C (50% RH) prior to heat acclimation | 38.9 | 153 | 52 pg·mL−1 (27%) s,c |
| Costa et al. [144] | 11 male (MT-HT) | 120 min running at 70% VO2max (fed) in Tamb 25 °C (35% RH) | N/A | 148 | 96 pg·mL−1 (46%) ns,c,# |
| Gill et al. [164] | 14 male and 3 female (HT) | 24 h ultramarathon (fed) in Tamb 0-20 °C (54–82% RH) | N/A | N/A | 122 pg·mL−1 (37%) s,# |
| Machado et al. (Part A) [174] | 9 male (MT) | 60 min running at 50% VO2max (fasted) in Tamb not reported | N/A | N/A | 130 pg·mL−1 (33%) ns,# |
| Machado et al. (Part B) [174] | 9 male (MT) | 60 min running at 50% VO2max (fasted) in Tamb not reported (FIO2 = 13.5%) | N/A | N/A | 250 pg·mL−1 (48%) s,# |
| Gaskell et al. [166] | 10 male and 8 female (MT-HT) | 120 min running at 60% VO2max (fed) in Tamb 35 °C (25% RH) | 38.6 | ~151 | LBP ~−2 µg·mL−1 (N/A%) ns,c |
| Selkirk et al. (Part A) [165] | 11 male (HT) | To fatigue (~163 min) uphill walk at 4.5 km.h−1 (fasted) in Tamb 40 °C (30% RH) | 39.1 | 164 | LBP ~0 µg·mL−1 (0%) ns |
| Moncada-Jiminez et al. [175] | 11 male (MT-HT) | 135-min laboratory duathlon at 71% VO2max (15km run and 30km cycle) (fasted) in Tamb not reported | 38.5 | N/A | LBP ~0.59 µg·mL−1 (22%) s,c |
| Selkirk et al. (Part B) [166] | 12 male (LT-MT) | To fatigue (~106 min) uphill walk at 4.5 km.h−1 (fasted) in Tamb 40 °C (30% RH) | 39.7 | 156 | LBP ~1.5 µg·mL−1 (15%) s |
| Jonvik et al. [140] | 16 male (HT) | 60 min cycling at 70% wattmax (fasted) in Tamb not reported | N/A | N/A | LBP 1.6 µg·mL−1 (13%) s |
| Costa et al. [144] | 11 male (MT-HT) | 120 min running at 70% VO2max (fed) in Tamb 25 °C (35% RH) | N/A | 148 | sCD14-ST 0.05 µg·mL−1 (N/A%) ns,c |
| Gaskell et al. [166] | 10 male and 8 female (MT-HT) | 120 min running at 60% VO2max (fed) in Tamb 35 °C (25% RH) | 38.6 | ~151 | sCD14-ST 0.1 µg·mL−1 (N/A%) s,c |
| Stuempfle et al. [158] | 15 male and 5 female (MT) | 161-km ultramarathon (26.8 ± 2.4 h; fed) in Tamb 0–30 °C (N/A RH) | 38.3 | N/A | sCD14-ST 0.6 µg·mL−1 (63%) s |
| Pugh et al. [134] | 10 male and 2 female (MT) | 42.4 km track marathon (4.1 ± 0.8 h; fed) in Tamb 16–17 °C (N/A RH) | N/A | ~160 | sCD14-ST 5.4 µg·mL−1 (164%) s,c |
| Nutrient | Evidence | Dosing | Consensus and Limitations |
|---|---|---|---|
| Carbohydrate | Cell: − − Clinical: + + − Exercise: + + + | 30–108 g·kg·h−1 liquid multi-transportable CHO. | Effects of pre- exercise CHO status or solid CHO ingestion unknown. Greater exploration on CHO timing and types required. |
| l-Glutamine | Cell: + + + − Clinical: + + − Exercise: + + + | 0.25–0.9 g·kg·FFM.−1 given 1–2 h pre-exercise. | Dose ≥ 0.25 g·kg·FFM−1 appears favourable. High doses poorly tolerated in some individuals. No evidence during prolonged exercise or on MT. |
| Bovine Colostrum | Cell: + + + + Clinical: + + + Exercise: + + | 20 g·day−1 for 14 days pre-exercise | Potentially useful following less demanding exercise. No effects with short-term supplementation. Certain formulations might be more beneficial. |
| Nitric Oxide | Cell: + + Clinical: + + Exercise: − − | More evidence required | No benefits of l-citrulline or sodium nitrate. Nitrate ingestion might compromise thermoregulation with exercise in the heat. Only two human exercise studies. |
| Probiotics | Cell: + − Clinical: + + − − Exercise: + − − | More evidence required | Contrasting results between formulations. Multi-strain probiotics seem favourable. Negative responses have been reported. Further evidence required. |
| Polyphenols | Cell: + + − − Clinical: + − Exercise: + − | 3 days of 0.5 g·day−1 of curcumin. Quercetin not recommended | Contrasting results between formulations. Only two human exercise studies. Further evidence required. |
| Zinc Carnosine | Cell: + + + Clinical: + + Exercise: + | 75 mg·day−1 for ≥ 2 days | Unknown effects in severe exercise situations. A 150 mg·day−1 dose warrants research. Only one human exercise study. Further evidence required. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ogden, H.B.; Child, R.B.; Fallowfield, J.L.; Delves, S.K.; Westwood, C.S.; Layden, J.D. The Gastrointestinal Exertional Heat Stroke Paradigm: Pathophysiology, Assessment, Severity, Aetiology and Nutritional Countermeasures. Nutrients 2020, 12, 537. https://doi.org/10.3390/nu12020537
Ogden HB, Child RB, Fallowfield JL, Delves SK, Westwood CS, Layden JD. The Gastrointestinal Exertional Heat Stroke Paradigm: Pathophysiology, Assessment, Severity, Aetiology and Nutritional Countermeasures. Nutrients. 2020; 12(2):537. https://doi.org/10.3390/nu12020537
Chicago/Turabian StyleOgden, Henry B., Robert B. Child, Joanne L. Fallowfield, Simon K. Delves, Caroline S. Westwood, and Joseph D. Layden. 2020. "The Gastrointestinal Exertional Heat Stroke Paradigm: Pathophysiology, Assessment, Severity, Aetiology and Nutritional Countermeasures" Nutrients 12, no. 2: 537. https://doi.org/10.3390/nu12020537